

Introduction
Materials and Methods
실험설계
실내초분광시스템
개발한 모델
전처리
모델 평가 및 실험 설정
온도 민감 대역 선정
주요 파장 선정
플로우차트
Results
온도별 스펙트럼 비교
개발된 모델 결과 비교
최종 모델
온도 민감 대역
최적 파장대역
Discussion
선행연구
한계점
향후연구
Conclusion
Introduction
복숭아(Prunus persica L. Batsch)는 전 세계적으로 중요한 온대 과수 중 하나로, 생과뿐만 아니라 주스, 잼, 통조림 등 다양한 가공식품의 원료로도 널리 활용되고 있다(Lurie and Crisosto, 2005). 특히 아시아, 유럽, 북미 등지에서 소비 수요가 높으며, 국내에서는 여름철 대표 과일로서 재배 면적과 생산량이 꾸준히 유지되고 있다(Arús et al., 2012). 이러한 경제적·문화적 중요성으로 인해 복숭아는 농가의 핵심 소득원으로 자리 잡고 있으며, 안정적인 생산과 품질 유지가 국가 및 지역 농업 전략에서 중요한 과제로 인식된다(Byrne et al., 2012).
그러나 복숭아의 안정적인 생산은 기상 조건에 크게 의존하며, 특히 개화기를 포함한 초기 생육 단계에서 온도 변화에 매우 민감하다(Grossman and DeJong, 1994). 시기에 발생하는 주요 위험 요인 중 하나가 봄철 서리이다(Li et al., 2012). 서리는 기온이 일시적으로 0 ℃ 이하로 내려가면서 대기 중 수증기가 결빙되거나 식물 표면의 수분이 얼어 조직 손상이나 고사를 유발하는 현상을 말한다(Byrne et al., 2012). 최근 이상기후로 인해 서리의 발생 빈도와 강도가 증가하고 있으며, 이러한 현상은 다양한 작물 중에서도 특히 과수에 치명적인 피해를 초래한다(Zhu et al., 2022). 복숭아 역시 예외가 아니며, 특히 개화기 전후의 저온 스트레스에 극도로 취약하다(Muthuramalingam et al., 2022). 짧은 기간의 급격한 기온 하강만으로도 꽃이 손상되어 착과율 및 최종 수확량이 크게 저하될 수 있다(Meng et al., 2009).
봄철 서리가 복숭아 생육과 수확에 미치는 영향에 대한 연구는 다수 수행되어 왔으나, 대부분 개화기 이후 착과 단계 또는 화학적 분석에 초점이 맞추어져 있다. 이는 개화 직전 단계의 꽃 조직에 대한 저온 스트레스 반응 및 피해 메커니즘에 관한 연구는 상대적으로 부족한 실정이다(Wang et al., 2013; Tanou et al., 2017; Zhang et al., 2023). 그러나 이 시기는 향후 개화 및 착과 성공 여부를 좌우하는 매우 민감한 시기로서, 피해를 조기에 진단하고 예방 기술을 마련하기 위해 보다 정밀한 연구가 필요하다(Penso et al., 2020).
초분광 이미징(hyperspectral imaging, HSI) 기술은 저온 스트레스에 따른 식물 조직의 미세한 생리·생화학적 변화를 비파괴적 방식으로 고해상도·고정밀도로 탐지할 수 있는 잠재력이 큰 방법론으로 주목받고 있다(Syeda et al., 2021). HSI는 수십에서 수백 개의 좁은 파장 대역(가시광선[visible, VIS]-근적외선[near-infrared, NIR], 400 - 1,000 nm 또는 확장된 단파적외선[short-wave infrared, SWIR] 대역)을 동시에 획득함으로써 각 픽셀마다 연속적인 스펙트럼 서명을 제공한다(Shahhosseini et al., 2021). 이를 통해 육안으로 식별하기 어려운 세포 구조 손상, 색소 조성 변화, 수분 함량 변동, 대사 이상 등을 스펙트럼 반사율 패턴으로 감지할 수 있다(Siche et al., 2016). 특히 실내 환경에서의 HSI 측정은 외부 환경 변수를 통제하고 균일한 광원 조건을 확보할 수 있어 잡음이 적고 신뢰도 높은 스펙트럼 데이터를 수집할 수 있다는 장점이 있다(Chen et al., 2021).
최근 HSI는 농업 분야에서 병해 조기 탐지, 수분·영양 상태 평가, 품질 등급 판별 등 다양한 연구에 적용되고 있으며, 이러한 기술적 특성은 개화 전 복숭아 꽃의 저온 스트레스 조기 탐지 및 피해 예측 모델 개발에도 응용될 가능성을 보여준다(Choi et al., 2025). 향후 HSI 기반 분석은 저온 피해에 취약한 시기의 정밀 모니터링과 함께, 인공지능(artificial intelligence, AI) 기반 데이터 해석 기술과 결합하여 복숭아 재배 관리 전략을 혁신적으로 개선할 수 있는 잠재력을 지닌다(Caballero et al., 2019).
초분광 데이터는 수백 개 이상의 파장 변수와 공간 정보를 포함하는 고차원 데이터로, 이를 효과적으로 해석하고 활용하기 위해서는 AI 기반의 첨단 분석 기법이 필수적이다(Wang et al., 2023a). 초분광 데이터는 비선형적이며 다중공선성이 존재하는 복잡한 특성을 가지므로, 단순한 통계적 접근만으로는 잠재 정보를 충분히 활용하기 어렵다(Alanazi et al., 2024). AI 기법은 이러한 복잡한 패턴과 변수 간 상호작용을 학습하여 저온 스트레스 여부 및 피해 정도를 조기에 평가하고 분류할 수 있는 가능성을 제공한다(Jiang et al., 2024).
또한 AI 기반 접근은 대규모 데이터에서 의미 있는 특징을 자동으로 추출하고, 사람의 육안이나 전통적 분석 방식으로는 포착하기 어려운 미세한 변화를 감지할 수 있다(Khonina et al., 2024). 이를 통해 개화 전 복숭아 꽃의 저온 피해를 실시간·정밀하게 진단하는 의사결정 지원 도구 개발로 확장될 수 있으며, 나아가 기후변화 대응형 스마트 농업 및 정밀농업 실현에도 기여할 수 있을것으로 판단한다.
따라서 본 연구에서는 개화 전 단계의 복숭아 중 천중도백도 꽃을 대상으로, 실내 환경에서 획득한 초분광 데이터를 활용하여 저온 스트레스에 따른 변화를 비파괴적으로 감지하고, 이를 바탕으로 머신러닝(machine learning, ML)과 딥러닝(deep learning, DL), 그리고 이들을 결합한 하이브리드 모델을 통해 온도 조건별 분류를 수행하고자 한다. 이러한 접근은 저온 피해를 조기에 탐지·예측할 수 있는 분석 기반을 마련하며, 향후 현장 적용이 가능한 정밀 모니터링 및 의사결정 지원 체계 개발에 기여하는 것을 목표로 한다. 본 연구의 구체적인 목적은 (1) 개화 전 복숭아 꽃의 초분광 스펙트럼 특성 분석, (2) 온도 조건별 데이터셋 구축과 적합한 전처리 기법 검토, (3) ML·DL·하이브리드 모델을 활용한 분류 성능 평가 및 비교를 통해 서리 피해 예측 가능성을 입증하는 것이다.
Materials and Methods
실험설계
본 연구에서는 개화 전 복숭아 꽃을 대상으로 총 네 가지 온도 조건(-4℃, -2℃, 0℃, 3℃)을 설정하여 저온 처리 실험을 수행하였다. 3℃처리 그룹은 2025년 6월 2일부터 6월 8일까지 개화 기간을 유지한 후, 6월 9일에 3℃에서 3시간, 8℃에서 3시간, 13℃에서 3시간의 순차적 온도 처리를 수행하였다. 0℃그룹 역시 동일한 개화 기간(6월 2일 - 6월 8일)을 거친 뒤, 6월 9일에 0℃에서 3시간, 6.5℃에서 3시간, 13℃에서 3시간의 처리를 실시하였다. -2℃그룹은 6월 3일부터 6월 9일까지 개화 기간을 유지하였으며, 6월 10일에 -2℃에서 3시간, 5.5℃에서 3시간, 13℃에서 3시간의 온도 처리를 적용하였다. -4℃그룹은 6월 8일부터 6월 10일까지 개화 기간을 유지한 후, 6월 11일에 -4℃에서 3시간, 4.5℃에서 3시간, 13℃에서 3시간의 저온 처리를 수행하였다. 모든 처리군에는 동일한 순차적 온도 처리 프로토콜이 적용되었다.
저온 처리는 각 개화 단계 종료 직후 수행되었으며, 처리 후 24시간 이내에 초분광 이미징 데이터를 수집하였다. 3℃및 0℃그룹은 온도 처리 다음 날인 6월 10일에 초분광 촬영을 실시하였고, -2℃그룹은 6월 10일 동일한 날에 저온 처리와 촬영을 모두 수행하였다. -4℃그룹은 6월 11일에 저온 처리 직후 초분광 데이터를 획득하였다. 모든 촬영 과정은 동일한 환경과 절차에 따라 수행되었으며, 각 온도 조건별 데이터 수집 시점은 처리 조건에 맞추어 표준화하였다(Table 1).
Table 1.
Temperature treatment conditions, flowering period, and hyperspectral image acquisition schedule for the four treatment groups.
초분광 이미징 촬영은 최적화된 설정값을 적용하여 수행하였다. 실험 수행을 위해 라인 개수는 680으로 설정하였으며, 파장 스텝 크기는 2.3 nm로 조정하였다. 반사광 모드에서의 노출시간은 40 µs으로 설정하였고, gain 값은 0으로 유지하였다. 스펙트럼 및 공간 샘플링은 각각 1로 설정하여 모든 스펙트럼 밴드 및 공간 픽셀을 원본 해상도로 획득하였다. 이후 노이즈 저감을 위해 프레임 평균값은 2로 설정하여 촬영을 진행하였다.
또한, 각 온도 처리 그룹별로 개화 전 시료 수를 기록하였다. 3℃그룹은 개화 전 112개의 시료를 수집하였으며, 0℃그룹은 개화 전 128개의 시료를 확보하였다. -2℃그룹은 개화 전 192개의 시료를 수집하였고, -4℃그룹은 개화 전 128개의 시료를 확보하였다. 모든 시료는 동일한 조건에서 처리 및 촬영되었으며, 온도 조건별·개화 단계별 데이터 확보를 위해 균일하게 표준화된 프로토콜을 적용하였다.
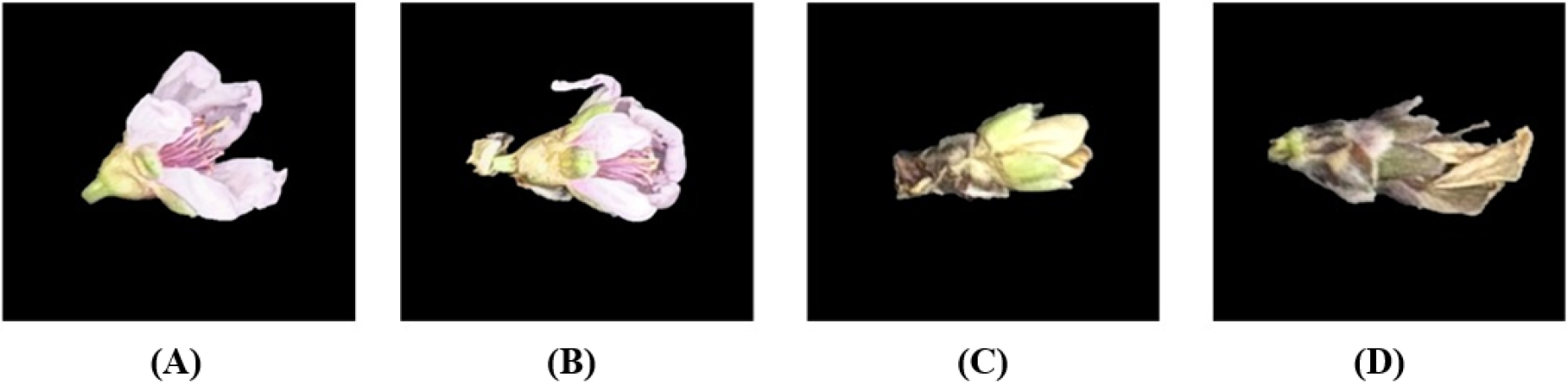
Fig. 1은 온도 조건에 따라 복숭아 꽃에서 나타난 냉해 수준의 차이를 시각적으로 비교한 것이다. (A) 3℃조건에서는 꽃잎과 꽃받침의 색이 선명하고 조직 손상이 관찰되지 않아 정상 생육 상태에 가까운 모습을 보인다. (B) 0℃에서는 시각적 변화가 미미하지만 일부 꽃잎 끝단에서 색이 연하고 수축된 형태가 부분적으로 확인되어 초기 냉해 반응이 나타난다. (C) -2℃에서는 꽃잎과 꽃받침 부위에서 갈변 및 수축이 명확히 발생하며 조직 괴사로 인한 변색이 뚜렷해 중등도 수준의 냉해 증상이 확인된다. 마지막으로 (D) -4℃에서는 전반적인 갈변과 왜곡, 심한 수축이 나타나며 꽃 전체가 고사한 형태를 띠어 고온 대비 냉해 피해가 극심한 상태임을 시사한다. 이러한 비교를 통해 온도 하강에 따라 복숭아 꽃 조직의 형태적 변형 및 색상 변화가 점진적으로 증가하며, 냉해 수준이 온도 의존적으로 심화됨을 확인할 수 있다.
실내초분광시스템
시스템 설계
본 연구에서는 식물 생육 데이터를 비파괴적으로 획득하기 위해 라인-스캔 기반 초분광 이미징 시스템을 구성·운용하였다. Fig. 2에서 확인할 수 있듯이, 시스템 프레임의 외형 치수는 가로 1,115 mm, 세로 516 mm, 높이 1,100 mm이며, 실내 촬영 시 외광 차단과 암실 형성을 위해 흑색 아크릴로 제작한 차광 커버(가로 1,164 mm, 세로 565 mm, 높이 1,164 mm)를 적용하였다. 차광 커버는 하부 개방형 박스 구조로 프레임 상부에 씌우는 방식이며, 전면 도어를 통해 내부 장비를 조작할 수 있도록 설계하였다. 계측 구성은 초분광 카메라(라인-스캔), RGB 카메라(FL3-U3-13E4C-C, FLIR, Canada), 할로겐 광원(LS-F100HSM-IR-FL, 100 W, 350 - 2,500 nm), 리니어 스테이지로 이루어졌다. VIS-NIR 촬영을 위해 광원 운용 대역은 400 - 1,000 nm로 설정하였고, 작업거리 330 mm에서 시료를 리니어 스테이지로 스텝 이송하여 라인-스캔 방식으로 초분광 데이터 큐브를 획득하였다.

시스템 보정
본 연구에서는 복숭아 꽃의 냉해 피해를 정량적으로 판별하기 위해, 초분광 영상 취득 이후의 데이터를 표준화된 규칙에 따라 자동 정리하고 분석 입력을 생성할 수 있는 전용 그래픽 사용자 인터페이스(graphical user interface, GUI)를 개발하였다(Fig. 3). 개발된 GUI는 초분광 기반 냉해 분석의 전 과정을 일관된 체계로 통합함으로써, 대량의 원시 영상을 효율적으로 처리하고 사용자 간 전처리 편차를 최소화하도록 설계되었다. 전체 구성은 입력 관리, 전처리 실행, 품질 검증의 세 가지 핵심 기능 영역으로 구분된다.
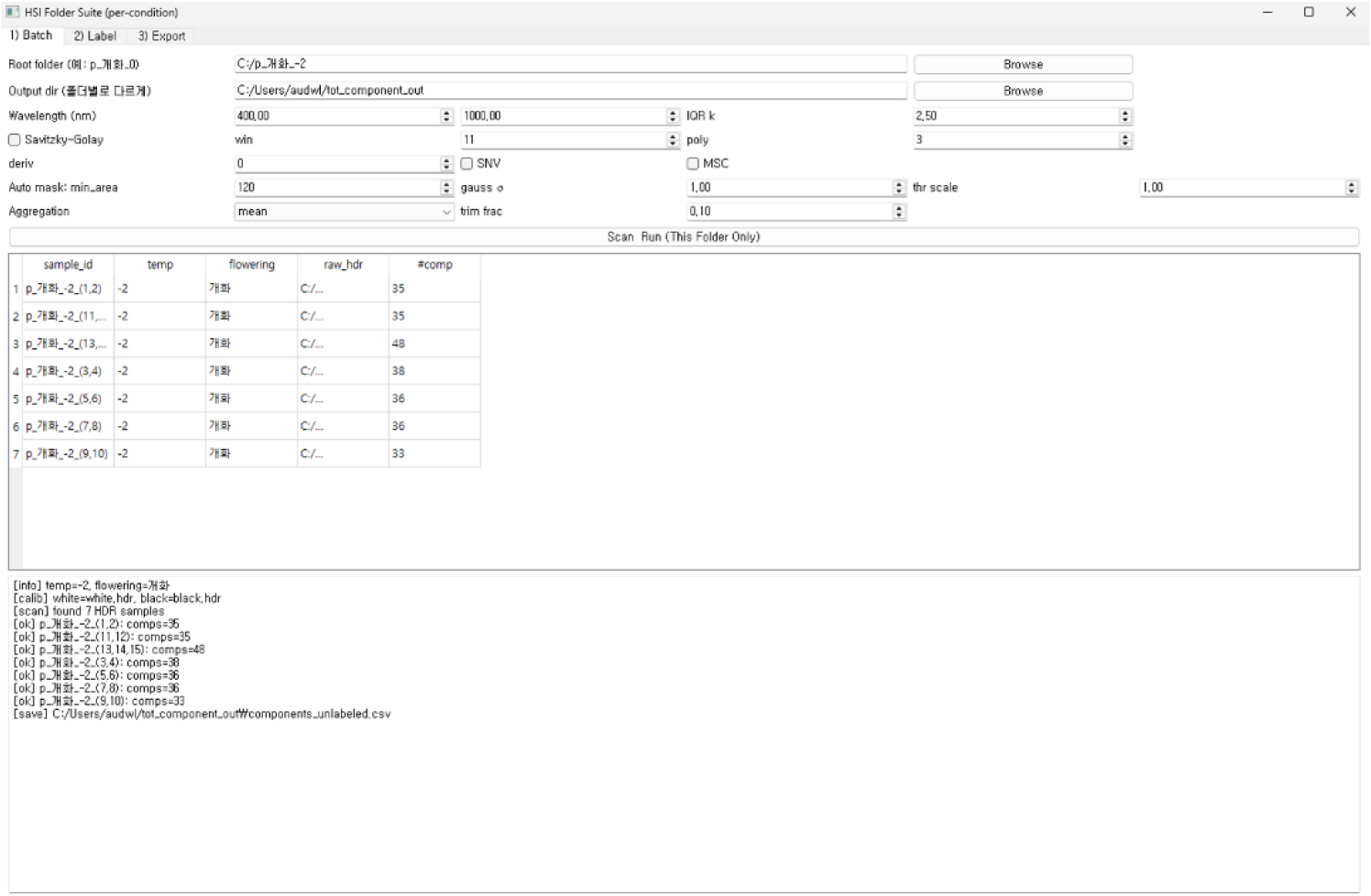
개발된 GUI 내에서 촬영된 ENVI(raw)·HDR 파일은 white 및 black 참조 영상과 함께 불러와 센서 특유의 밝기 변화를 보정하고, 파장축 정렬 및 밴드 간 간격 보정 과정을 통해 분석용 스펙트럼으로 변환하였다. 반사광 모드에서 획득된 원시 스펙트럼은 white/black 보정을 통해 상대 반사율 형태로 변환되었으며, Savitzky-Golay 필터링, 스펙트럼 미분, multiplicative scatter correction (MSC), standard normal variate (SNV) 등 스펙트럼 기반 전처리 기법을 적용해 잡음과 산란 효과를 최소화하였다. 또한 개화 개체 단위의 ROI를 기준으로 대표 스펙트럼을 산출하여 조건 간 비교가 가능한 표준화된 스펙트럼 테이블을 구축하였으며, HSI 데이터에서 하나의 꽃에서 복수의 ROI가 생성될 수 있는 점을 고려하여 데이터 분할은 ROI가 아닌 개체(꽃) 단위로 수행하였다. 이러한 일련의 캘리브레이션 및 전처리 과정은 냉해 조건에 따른 주요 파장대 차이를 정확하게 반영하고, 후속 모델링 단계에서의 분석 재현성을 확보하기 위한 기반으로 활용되었다.
본 GUI는 초분광 영상과 실험 지표를 기반으로 냉해 피해 분석에 필요한 데이터 입력, 전처리, 품질 검증, 분석용 출력 생성까지의 전 과정을 단일 플랫폼에서 통합적으로 제공한다. 특히 동일 규칙의 전처리를 적용함으로써 조건 간 비교 가능성과 분석 공정성이 확보되었으며, 주요 파장대 탐지 및 스펙트럼 기반 시각 자료까지 함께 제공함으로써 모델 해석력을 강화하였다. 이러한 구성은 대규모 초분광 데이터를 효율적·일관적으로 처리할 수 있는 기반을 제공하며, 복숭아 꽃 냉해 진단 연구의 정확성과 재현성을 동시에 향상시키는 역할을 한다.
개발한 모델
ML
본 연구에서는 복숭아 꽃을 대상으로 네 가지 온도 조건에서 분류하기 위해 ML모델인 RF (random forest), SVM (support vector machine) 및 MLP (multilayer perceptron) 기반 분류 모델을 구축하였다. 추가적으로 RF, ExtraTrees, SVM을 소프트 보팅(VotingClassifier)으로 결합시킨 앙상블 모델을 설계하였다.
RF는 다수의 의사결정나무를 독립적으로 학습시킨 뒤 다수결 투표 방식으로 최종 예측을 수행하여 모델의 편향을 줄이고 분산을 안정화한다(Saleem et al., 2021). 이 과정에서 과적합을 효과적으로 방지하며, 입력 변수의 상대적 중요도를 정량적으로 산출할 수 있어 해석 가능성이 높다(Ray et al., 2023).
SVM은 데이터를 고차원 특징 공간으로 사상하여 최대 마진을 갖는 초평면을 탐색함으로써 클래스 간 분리를 최적화한다(Bansal et al., 2022). 커널 함수를 활용하면 비선형 경계 학습이 가능하므로 복잡한 패턴이 포함된 온도별 생리 반응 데이터 분류에도 적합하다(Wang et al., 2023b).
MLP는 입력층, 복수의 은닉층, 출력층으로 구성된 심층 신경망으로, 활성화 함수를 통해 비선형 변환을 수행하며 데이터의 복잡한 관계를 학습한다(Iqbal et al., 2024). 역전파 알고리즘을 이용해 가중치를 반복적으로 최적화하고, 은닉층의 개수, 뉴런 수, 학습률 등 다양한 하이퍼파라미터 조합을 통해 모델의 표현력과 일반화 성능을 조절할 수 있다(Bazrafshan et al., 2022).
VotingClassifier은 여러 개의 이질적인 분류기를 결합하여 예측 확률을 평균한 뒤 최종 출력을 결정하는 앙상블 학습 기법이다. 개별 분류기의 장점을 통합함으로써 단일 모델 대비 편향과 분산을 줄일 수 있으며, 보다 안정적이고 강건한 예측 성능을 확보할 수 있다(Khatun et al., 2023). 본 연구에서는 RF, ExtraTrees, SVM을 결합하여 소프트 보팅 모델을 구성하였으며, 이를 통해 복숭아 꽃 온도 분류 문제에서 일반화 성능 향상을 도모하였다.
본 연구에서는 동일한 복숭아 꽃 온도별 스펙트럼 데이터셋을 활용하여 개발된 모델들을 비교·평가하고, 가장 적합한 분류 모델을 도출하였다. 또한, 본 연구에서는 데이터의 일반화 성능을 확보하기 위해 전체 데이터셋을 80% 학습용, 20% 테스트용으로 분할하였으며, 모델 성능의 안정성을 확보하기 위해 k-fold 교차검증을 수행하였다.
DL
본 연구에서는 동일한 데이터셋을 활용하여 세 가지 형태의 DL도 구축하였다. DL은 대규모 데이터로부터 비선형적 패턴을 자동으로 학습할 수 있는 기법으로, 특히 고차원 영상·스펙트럼 데이터의 복잡한 특징을 효과적으로 추출하는 데 강점을 가진다(Dolaptsis et al., 2024). 초분광 데이터는 파장별 연속적 스펙트럼 정보를 포함하고 있어, 이러한 DL 기반 특징 학습 방식과 높은 궁합을 보이며 정밀한 분류 성능을 구현할 수 있다(Nikzadfar et al., 2024). 특히 CNN (convolutional neural network)은 합성곱 연산을 통해 입력 데이터의 지역적·계층적 특징을 효율적으로 추출하는 구조로, 초분광 데이터의 파장 축에서 발생하는 미세한 변화 패턴을 효과적으로 학습할 수 있다는 장점을 가진다(Gavahi et al., 2021).
1D CNN은 개별 스펙트럼 데이터를 입력으로 받아 1차원 합성곱 연산을 통해 파장 축에서의 지역적 특징을 학습하였다. 2D CNN은 ROI에서 추출한 2차원 스펙트럼 이미지를 입력으로 하여 공간적 패턴과 스펙트럼 간 상호작용을 동시에 고려하였다. 하이브리드 1D-2D CNN은 스펙트럼 기반 1D 특징과 이미지 기반 2D 특징을 병합하여 최종 분류층에 전달함으로써 두 입력의 상호 보완적 정보를 활용하였다.
이러한 세 가지 DL 모델은 서로 다른 수준의 특징 추출 능력을 제공하며, 복숭아 꽃의 온도별 피해 반응을 보다 정밀하게 분류하기 위해 상호 비교·평가되었다. ML과 마찬가지로, 전체 데이터셋은 80%를 학습용, 20%를 테스트용으로 분할하였으며, 모델의 성능 안정성을 확보하고 과적합을 방지하기 위해 k-fold 교차검증을 수행하였다.
전처리
본 연구는 초분광 스펙트럼 전처리 방법의 효과를 독립적으로 비교하기 위해 Normalization, MSC, SNV, interquartile range (IQR) 기반 이상치 제거를 각각 단일 조건으로 구성하여 동일한 원시 데이터, 분류기 및 하이퍼파라미터 환경에서 전처리만 교체하여 5-fold 교차검증을 수행하였다. 모든 변환에 필요한 통계량은 각 fold의 훈련 데이터에서만 추정하여 데이터 누수를 방지하였다. 또한 스펙트럼 평활화를 위해 Savitzky-Golay 필터(window length = 11, polynomial order = 2, derivative order = 0)를 적용하였다.
Normalization은 밴드 간 스케일 불일치를 완화하여 모델 수렴 안정성과 특성 간 비교 가능성을 향상시켰다(Witteveen et al., 2022). MSC는 훈련 데이터의 평균 스펙트럼을 기준으로 하여 각 스펙트럼의 가산 산란과 곱셈 산란 성분을 보정함으로써 표면 상태, 조명 및 시료 두께 변화에 따른 산란 효과를 완화하였다(Li et al., 2021). SNV는 각 스펙트럼을 평균 중심화한 후 표준편차로 정규화하는 방식으로 적용하여 시료 간 산란 편차를 보정하였다(Zhang and Abdulla, 2023). IQR 기반 이상치 제거는 전체 스펙트럼 벡터를 기준으로 계산된 분포에서 Q1 − 1.5 × IQR 및 Q3 + 1.5 × IQR 범위를 벗어나는 시료를 이상치로 판단하여 제거하는 방식으로 수행되었다(Choi et al., 2026).
전반적으로 전처리 방법의 상대적 효과는 데이터 교란의 주요 원인인 스케일 불일치, 산란 및 잡음, 시료 간 산란 편차, 그리고 이상치 혼입의 정도에 따라 달라질 수 있으며, 본 비교 설계는 현장 적용 시 단순하고 강건한 전처리 방법을 선택할 수 있도록 한다.
모델 평가 및 실험 설정
개발된 모델의 성능 평가는 테스트 데이터셋을 이용하여 수행하였다. 평가는 모델 예측값과 실제 정답을 비교하는 혼동 행렬(confusion matrix)을 기반으로 이루어졌으며, 혼동 행렬은 참양성(true positive, TP), 참음성(true negative, TN), 거짓양성(false positive, FP), 거짓음성(false negative, FN)으로 구성된다. 혼동 행렬을 바탕으로 각 모델의 분류 성능을 정량적으로 평가하기 위해 정확도(accuracy), 재현율(recall), 정밀도(precision), F1-점수(F1-score)와 같은 주요 성능 지표를 산출하였다. 각 성능 지표의 산출식은 다음과 같다.
데이터셋의 크기 제한을 보완하기 위해 5-겹 교차검증을 적용하였다. 클래스 불균형으로 인한 평가 편향을 완화하기 위해 각 fold가 원본 클래스 분포를 유지하도록 층화 식을 적용하였으며, 매 분할 시 데이터는 무작위로 섞였고 재현성을 확보하기 위해 고정 난수 시드(random_state = 42)를 사용하였다. 각 fold에서 전체 데이터의 약 80%는 학습용, 20%는 검증용으로 할당되었다. 모든 전처리 과정(예: 특성 스케일링)은 학습용 데이터에 대해서만 수행되었고, 이후 동일한 변환을 해당 검증 데이터에 적용하여 데이터 누수를 방지하였다. 공정한 비교를 위해 동일한 fold 분할이 모든 모델에 일관되게 적용되었다. 정확도, macro 및 weighted F1-점수, 정밀도, 재현율 등 성능 지표는 5개 fold의 평균값과 표준편차(mean ± standard deviation)로 보고하였다.
모든 실험은 Windows 운영체제 환경에서 Jupyter Notebook (Anaconda Inc., USA)을 이용하여 수행하였다. 모델 학습 및 추론에는 GeForce GTX 1060 GPU (NVIDIA, USA)를 사용하였다. 프로그래밍 환경은 Python 3.9.18 (Python Software Foundation, USA)이었으며, DL 프레임워크로는 TensorFlow 2.10.0 (Google Brain Team, USA)을 사용하였다.
온도 민감 대역 선정
각 파장대에서 온도 요인이 반사도 변화에 미치는 영향을 정량적으로 평가하기 위해 η2 (effect size) 분석을 수행하였다(Brown, 2008). η2은 0 - 1 범위를 가지며, 값이 클수록 온도 처리가 해당 파장에서의 반사도 변동을 더 잘 설명함을 의미한다(Richardson, 2011). 본 연구에서는 각 파장 λ마다 온도 처리에 의한 제곱합(SS_between)과 총 제곱합(SS_total)을 계산하여 η2 = SS_between / SS_total로 정의하였다. 산출된 η2 값은 스펙트럼 파장대별로 배열하고 라인 프로파일 형태로 시각화하여 온도 반응 민감도가 높은 파장 구간을 도출하였다(Li et al., 2023). 이러한 방식은 온도 요인의 효과 크기를 비모수적으로 해석할 수 있으며, 특정 스펙트럼 대역이 저온 반응에 기여하는 정도를 정량적으로 비교하는 데 유용하다.
주요 파장 선정
본 연구에서는 복숭아 꽃의 온도 분류 모델이 실제로 어떤 파장 정보를 기반으로 예측을 수행하는지를 규명하기 위해 적분 기울기(integrated gradients, IG) 기법을 적용하였다. IG는 기준 입력에서 실제 입력으로 변환되는 경로를 따라 기울기를 적분함으로써, 입력 스펙트럼의 각 파장이 모델의 예측에 기여하는 정도를 정량화하는 해석적 기법이다(Walker et al., 2024). 본 연구에서는 기준 스펙트럼(baseline)을 평균 반사도 스펙트럼으로 설정하고, IG 점수를 통해 파장별 중요도를 산출하였다. 계산된 중요도는 정규화 후 파장 축에 매핑하여 시각화하였으며, 이를 통해 모델이 분류 과정에서 활용한 주요 파장 구간을 식별하였다.
플로우차트
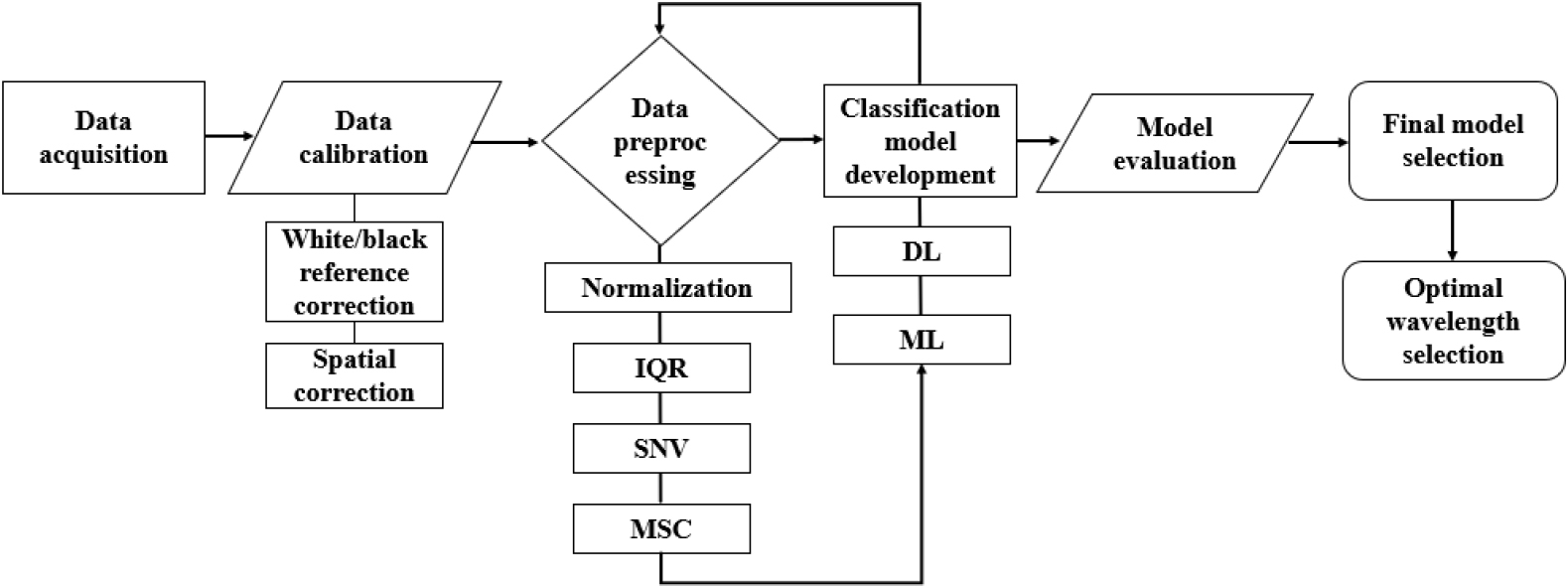
본 연구의 전체 분석 절차는 Fig. 4에 요약되어 있다. 먼저 실험 샘플의 영상을 획득한 뒤, 화이트·다크 레퍼런스를 활용하여 기초 보정 과정을 수행함으로써 촬영 환경에 따른 밝기 편차와 측정 오차를 최소화하였다. 보정된 데이터는 전처리 단계로 전달되었으며, 스펙트럼 품질 향상과 모델 입력의 일관성을 확보하기 위해 정규화 및 기본적인 스펙트럼 교정 기법을 적용하였다. 이러한 전처리 과정은 전체 데이터의 변동성을 줄이고 이후 단계에서의 안정적인 학습 기반을 마련하는 데 목적이 있다.
전처리가 완료된 데이터는 ML과 DL 기반의 분류 모델 구축에 사용되었다. 두 접근법은 동일한 데이터셋을 바탕으로 독립적으로 학습 및 검증되었으며, 모델 구조의 특성과 입력 형식에 따라 스펙트럼 정보를 해석하는 방식이 달라 다양한 관점에서 성능을 비교할 수 있도록 구성하였다. 학습된 모델들은 동일한 평가 지표를 활용하여 성능을 정량적으로 분석하였고, 그 결과를 토대로 최종적으로 가장 우수한 분류 모델을 선정하였다.
마지막으로 선정된 모델을 기반으로 주요 파장 정보를 확인하여 모델이 활용한 특징의 해석력을 높이고, 초분광 데이터가 제공하는 분광적 민감성이 실제 분류 과정에 어떻게 기여했는지를 검토하였다. 전체적으로, 본 연구의 분석 파이프라인은 데이터 획득부터 보정·전처리, 모델링, 평가, 그리고 파장 해석에 이르는 일련의 흐름을 체계적으로 구성하여 모델의 신뢰성과 적용 가능성을 확보하는 것을 목표로 하였다.
Results
온도별 스펙트럼 비교
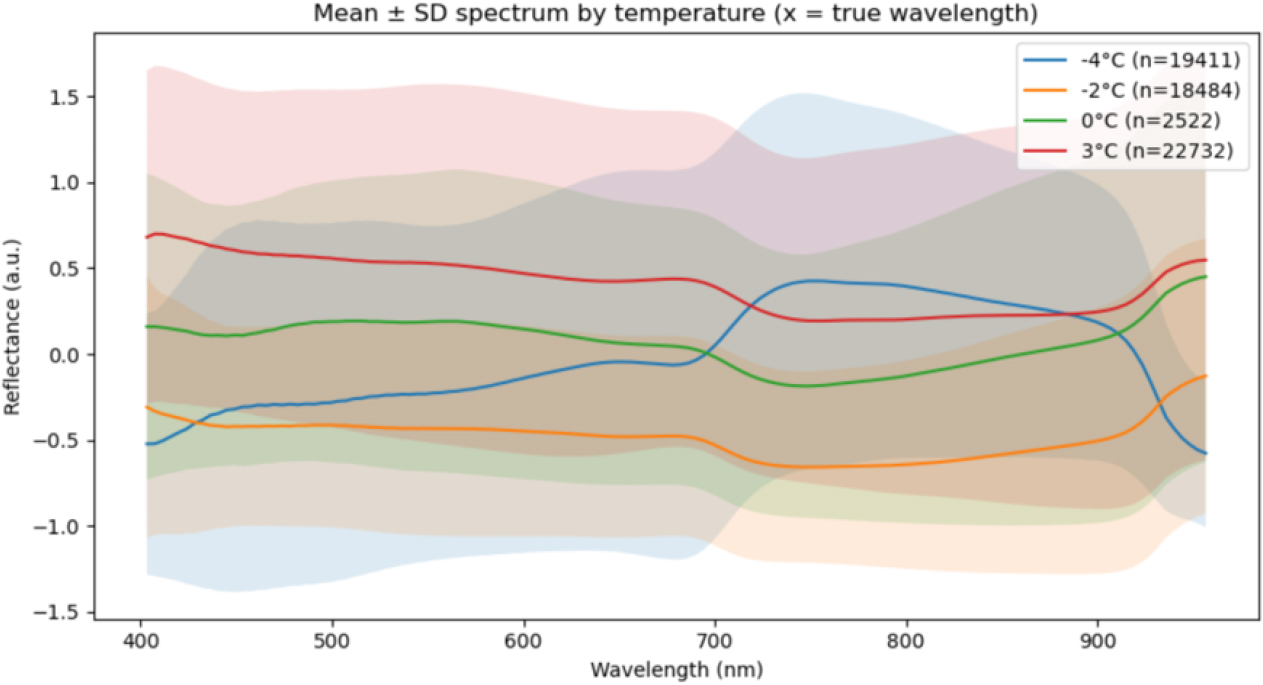
Fig. 5는 복숭아 꽃의 온도 조건별 평균 ± 표준편차 스펙트럼을 나타낸 것이다. 가로축은 파장, 세로축은 반사율이며, 음영 구간은 파장별 반사도의 표준편차를 의미한다. 전반적으로 온도가 낮을수록 반사율이 감소하는 경향을 보였으며, 이는 세포 구조 손상 및 수분 함량 저하에 따른 산란 강도 감소로 해석된다.
-4℃조건에서는 전 구간에서 반사율이 가장 낮고, 특히 800 nm 이후 근적외선(near-infrared, NIR) 영역에서 급격히 상승하는 형태를 나타냈다. -2℃에서는 -4℃보다 반사율이 다소 높으나 여전히 낮은 수준을 유지하였고, 700 - 900 nm 구간에서 변동 폭이 크게 나타났다. 한편, 3℃조건은 정상 상태의 대조군으로, 전 파장 영역에서 가장 안정된 반사율 곡선을 나타냈다. 이 구간에서는 553 nm와 850 - 900 nm에서 반사율이 안정적으로 유지되었으며, 이는 광합성 색소가 정상적으로 유지되고 세포 구조가 안정된 건강한 상태를 의미한다.
0℃조건의 스펙트럼은 3℃와 형태적으로 유사하였으며, 주요 전환점에서도 큰 차이가 관찰되지 않았다. 이는 경계 온도에서 스트레스 반응이 미약하거나 초기 단계에 해당함을 의미한다. 결과적으로, 553 nm와 850 - 900 nm가 온도에 따른 스펙트럼 차이를 가장 뚜렷하게 구분하는 핵심 파장대로 확인되었다. 이는 정상 상태(3℃) 대비 온도가 낮아질수록 반사율이 감소하고, 저온 스트레스가 진행됨에 따라 광합성 색소 및 세포 구조의 변형이 반사 스펙트럼 형태로 명확히 나타남을 의미한다.
개발된 모델 결과 비교
본 연구에서는 복숭아 꽃의 냉해 피해를 온도 수준에 따라 정량적으로 평가하기 위해 총 일곱 개의 분류 모델을 구축하였다. 이 중 네 개는 ML 기반 모델이고 세 개는 DL 기반 모델로 구성되었다. 네 개 온도 조건(-4℃, -2℃, 0℃, 3℃)에 대한 분류 결과는 Table 2에 제시되어 있으며, IQR 전처리를 적용한 1D CNN이 가장 높은 정확도를 나타냈다. 해당 모델의 혼동행렬 및 성능 지표는 Fig. 6에 제시되어 있다.
Table 2.
Accuracy results of the four-temperature classification models.
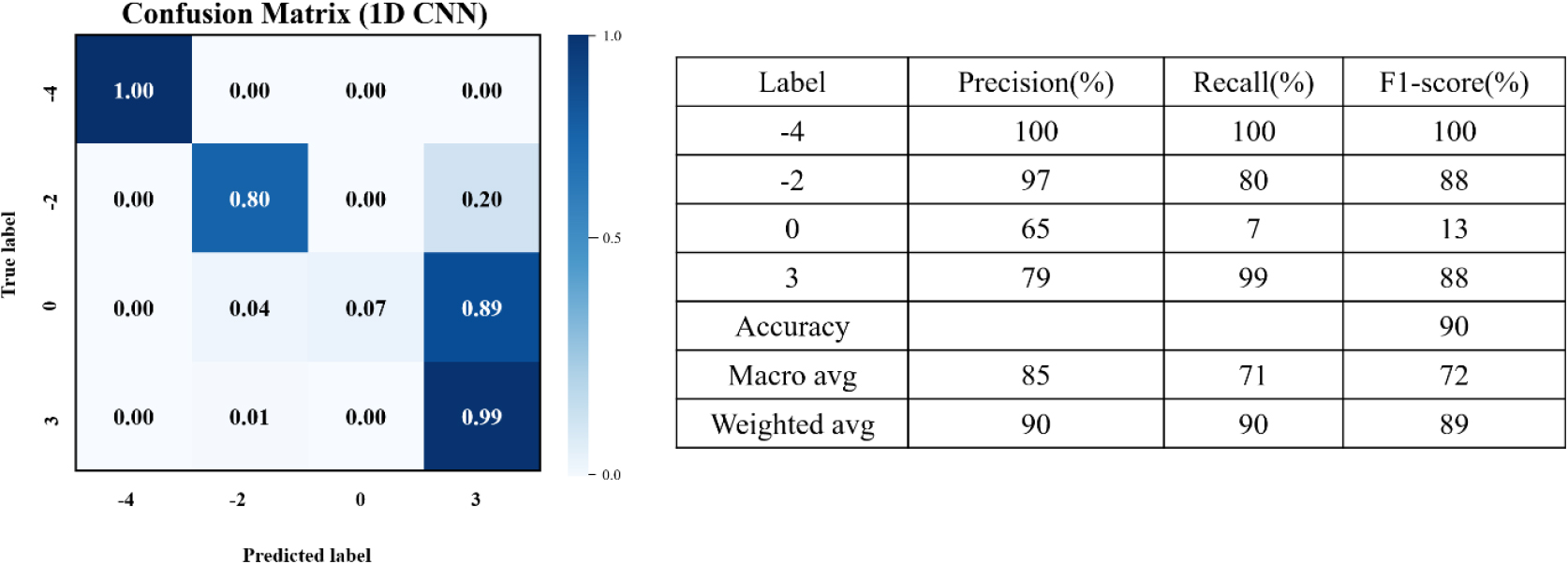
4개 온도 조건을 동시에 분류한 결과, Fig. 7에서 확인되듯이 -4℃와 3℃는 비교적 명확하게 구분되었으나, 0℃는 3℃와 혼동되는 경향을 보였다. 이는 냉해 발생의 경계 온도 영역에서 조직 손상이 부분적으로 발생하거나 아직 명확히 진행되지 않은 상태가 혼재되어 있기 때문으로 판단된다. 이러한 상태에서는 스펙트럼 반응이 정상 조직과 유사하게 나타날 수 있어 분광 패턴의 중첩이 발생할 가능성이 있다.
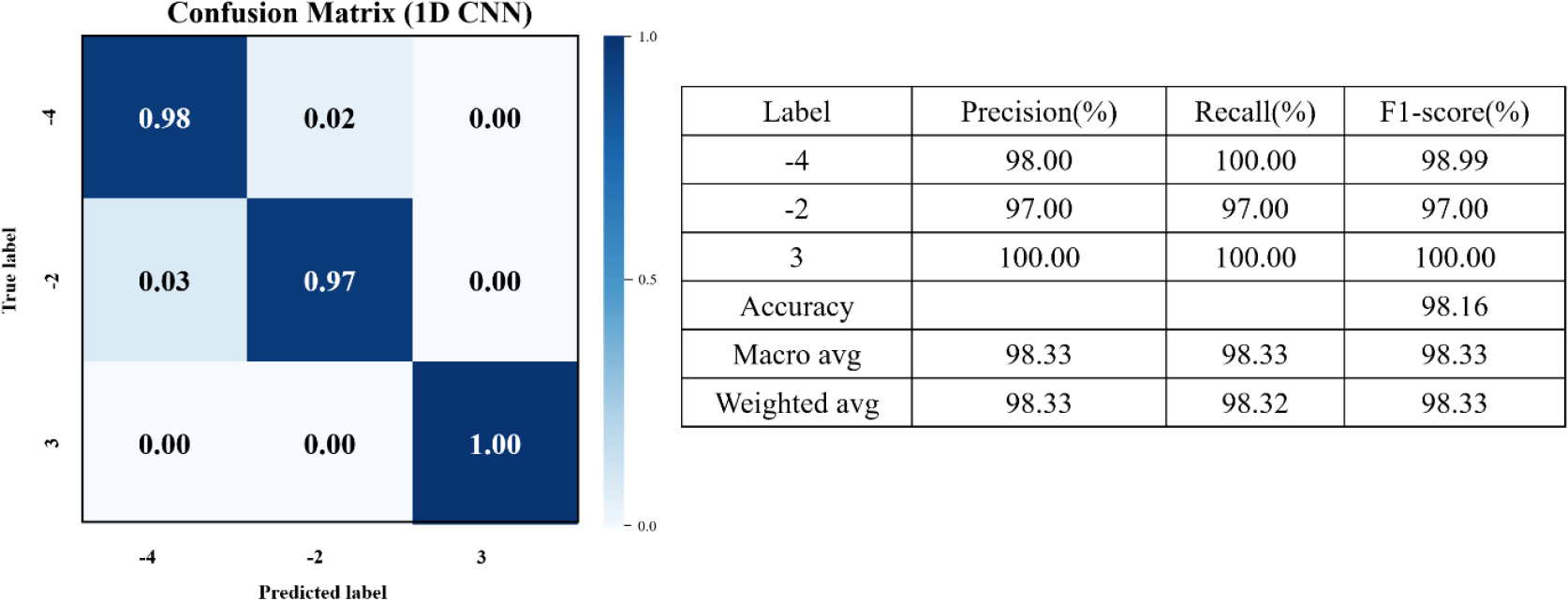
경계 온도 구간에서 발생하는 이러한 분광 중첩 현상이 분류 성능에 미치는 영향을 보다 명확히 분석하기 위해, 본 연구에서는 0℃ 클래스를 제외한 세 개 온도(-4℃, -2℃, 3℃)를 대상으로 추가적인 분류 분석을 수행하였다. Table 3의 결과에 따르면 1D CNN은 대부분의 전처리 조건에서 여전히 가장 우수한 성능을 보였으며, 전체 정확도 또한 4개 클래스 분류 결과 대비 향상되는 경향을 나타냈다. 이는 경계 온도 영역에서 나타나는 스펙트럼 유사성이 다중 분류 과정에서 주요 혼동 요인으로 작용했음을 시사한다. 예시적으로, IQR 전처리를 적용한 1D CNN의 세 개 온도 분류 성능은 Fig. 7에 나타나 있으며, 모든 클래스에서 97 - 100%의 높은 정분류율을 보였다.
Table 3.
Accuracy results of the three-temperature classification model.
한편 실제 농업 현장에서의 냉해 진단은 모든 온도 수준을 세분화하여 판별하기보다는, 피해 발생 여부를 신속하게 판단하는 의사결정 형태로 활용되는 경우가 많다. 이러한 적용 관점을 고려하여 본 연구에서는 심각한 냉해 피해가 발생한 조건(-4℃)과 정상 생육 조건(3℃)을 구분하는 이진 분류 모델을 추가적으로 구축하였다. 이러한 이진 분류 분석은 모델의 실제 현장 적용 가능성을 평가하기 위한 목적으로 수행되었다.
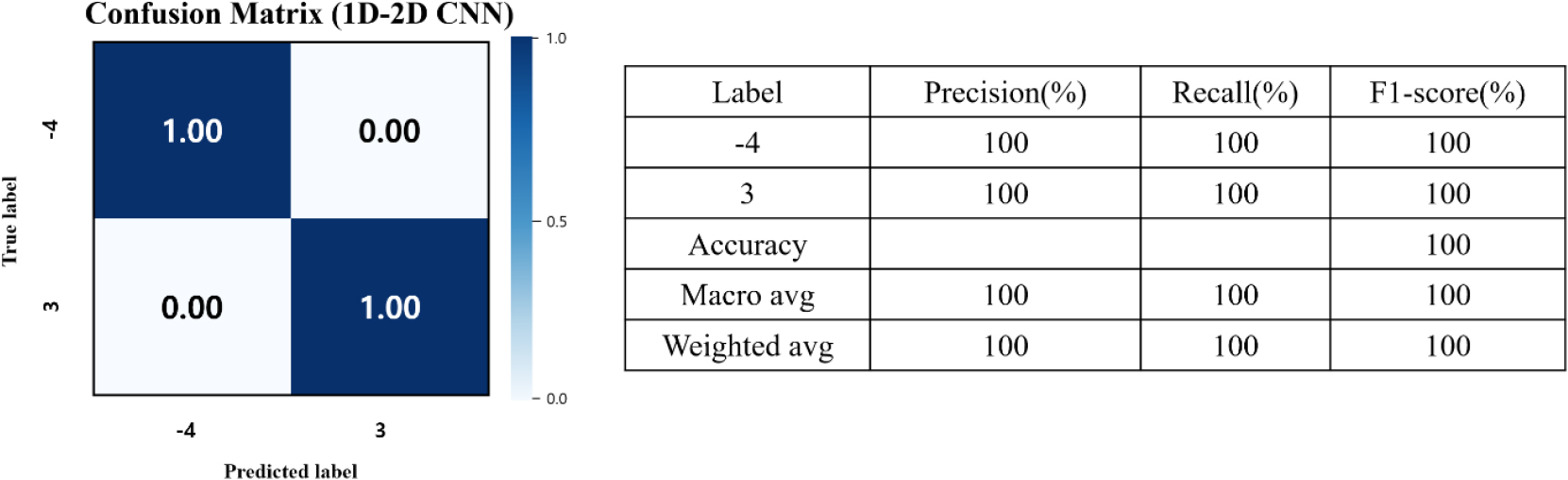
최종 이진 분류 결과는 Table 4에 제시되어 있으며, 하이브리드 1D-2D CNN 모델이 정확도 1.00을 달성하여 모든 모델 중 가장 우수한 성능을 보였다. 또한 IQR 전처리를 적용한 1D-2D CNN의 혼동행렬 및 전체 성능 지표는 Fig. 8에 제시되어 있으며, 모든 평가 지표에서 100%를 기록하였다. 이는 1D 스펙트럼 특징과 2D 공간적 정보를 동시에 활용하는 멀티모달 구조가 온도 변화에 따른 분광 및 형태학적 차이를 효과적으로 반영했기 때문으로 판단된다.
Table 4.
Accuracy results of the two-temperature classification model.
최종 모델
전체적으로 IQR 전처리를 사용한 CNN 모델이 전 모델에서 가장 우수한 성능을 나타냈다. 전체적으로 IQR 전처리를 적용한 CNN 계열 모델이 복숭아 꽃 온도 분류에서 가장 우수한 성능을 보였다. 이는 단순히 모델의 구조적 특성 때문이 아니라, 스펙트럼 데이터의 통계적 분포와 광학적 특성이 IQR과 CNN의 결합에 적합했기 때문이다.
현장 스펙트럼은 조명 세기, 표면 반사, 배경 간섭 등으로 인해 이상치가 빈번히 발생하며, 이는 모델 학습 시 분산 왜곡과 경계 불안정을 초래한다(Syu et al., 2019). IQR기반 전처리는 이러한 극단값을 제거해 데이터 분포의 중심부(중앙 50%)를 안정화하고, 파장별 반사율의 변동 폭을 완화시켰다(Gowthami and Priscila, 2024). 이로써 클래스 간 평균 차가 명확해지고, 경계 온도(0℃, -2℃) 구간에서도 클래스 간 중첩이 감소하였다. 또한, IQR은 표준화나 정규화 대비 형태적 정보를 왜곡하지 않아, 이후 CNN이 파장 축의 연속적 패턴을 효율적으로 학습할 수 있도록 했다.
CNN 모델은 스펙트럼의 연속적 형태 정보를 자동으로 학습할 수 있는 구조적 이점을 가진다(Alzubaidi et al., 2021). 1D CNN은 파장 축을 따라 흡수 피크(≈680 nm), shoulder (≈553 nm), red-edge (700 - 750 nm), NIR 반사(≥ 850 nm) 등의 특징을 합성곱 필터로 포착하여 비선형적 상호작용을 다층적으로 학습한다(Qazi et al., 2022). 이러한 방식은 고정 특징을 사용하는 ML 모델에 비해 경계 온도에서의 미세한 반사율 차이를 효과적으로 구분할 수 있도록 한다(Shirazi et al., 2024). 또한, CNN의 파라미터 공유와 지역 필터 구조는 스펙트럼 간의 공선성 문제를 완화하고, 효율적인 차원 축소 및 규제 효과를 제공하여 과적합을 줄인다(Ige and Sibiya, 2024). 특히 하이브리드 1D-2D CNN 구조는 1D 스펙트럼에서 화학적·수분 관련 신호를, 2D 영상에서 조직·형태적 정보를 동시에 학습한다(Li et al., 2019). 이러한 멀티모달 융합은 -2℃와 3℃처럼 NIR 스펙트럼이 유사한 클래스 간 혼선을 완화하며, 결정 경계의 구분성을 크게 향상시켰다(Cai and Chen, 2020).
IQR 전처리로 안정화된 입력을 바탕으로 CNN이 형태 기반 핵심 신호를 다중 스케일로 학습함으로써 경계 온도에서의 오분류를 최소화하였고, 이러한 IQR-CNN 결합은 스펙트럼 분포 안정화와 형태 학습의 상호보완적 효과를 통해 복숭아 꽃 온도 구분 정확도를 향상시켜 냉해 감지 및 온도 반응형 모니터링 모델의 핵심 기반이 될 수 있음을 시사한다.
온도 민감 대역
Fig. 9에서 확인할 수 있듯이 전체적으로 η2 값은 0.10 - 0.30 범위에 분포하여, 전 파장 영역에서 온도가 일정 수준 이상 반사율에 영향을 미치지만 일부 특정 구간에서 특히 큰 반응이 나타남을 확인할 수 있었다.
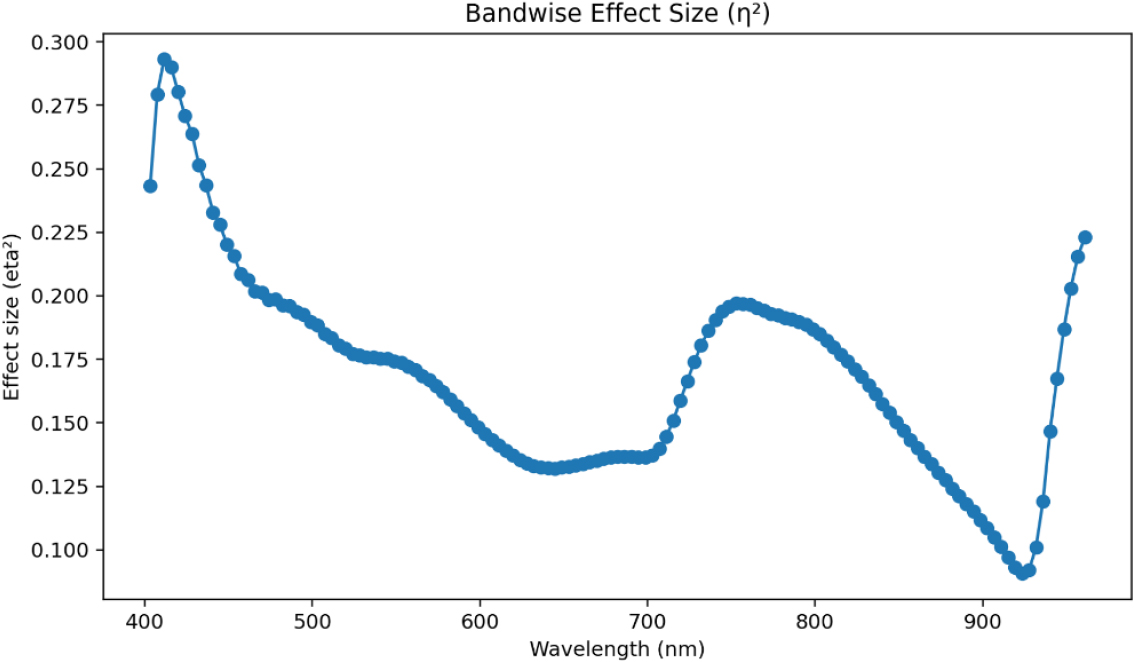
먼저 400 - 420 nm 구간에서 η2 값이 약 0.28 - 0.30으로 가장 높게 나타났다. 이 구간은 엽록소와 안토시아닌 등 광합성 색소의 흡수대에 해당하며, 저온 노출 시 색소 농도 및 조성 변화가 즉각적으로 반영된 결과로 해석된다(Thron et al., 2001). 즉, 냉해 환경에서 색소의 광흡수 능력이 변화함에 따라 가시광 반사율의 변동이 크게 나타난 것이다.
700 - 800 nm 구간은 red-edge 전이 영역으로, η2 값이 약 0.19 - 0.20 수준을 보여 중간 정도의 온도 민감도를 나타냈다(Mutanga and Skidmore, 2007). 이는 광합성 조직의 구조적 변화나 수분 함량 저하 등 세포 수준의 조직 반응이 반영된 결과로 판단된다(Gitelson et al., 1996). 또한 이 구간은 식물 조직 내 산란과 흡수가 동시에 영향을 미치는 파장대로, 저온 조건에서의 세포벽 파손이나 기공 구조 변화가 반사 신호를 변화시킨 것으로 보인다(Gupta et al., 2003).
한편 900 nm 이후 근적외선(NIR) 영역에서는 η2 값이 다시 약 0.22까지 상승하여, 온도 조건에 따른 세포 구조 및 수분 상태의 차이가 뚜렷하게 반영된 것으로 나타났다. 이 영역은 세포 내 자유수의 흡수 특성과 관련이 있으며, 냉해로 인한 세포 탈수와 수분 재분포가 반사율에 영향을 준 것으로 해석된다(Peng et al., 2021).
요약하면, η2 분석 결과 400 - 420 nm (색소 반응대), 700 - 800 nm (red-edge), 900 nm 이후에서 온도 민감도가 높게 나타났다. 이는 냉해 조건에서 이러한 파장대가 온도 변화에 민감하게 반응하는 핵심 영역임을 보여주며, 향후 냉해 피해 조기 진단 및 온도 반응형 모니터링의 주요 파장 후보로 활용될 수 있을 것으로 판단된다.
최적 파장대역
각 최종 모델에서 예측 기여도가 높은 상위 5개 중요 파장대를 도출하였다. 4개 온도 분류(-4℃, -2℃, 0℃, 3℃)에서는 IQR 전처리를 적용한 1D CNN 모델, 3개 온도 분류에서는 IQR 및 정규화 전처리를 병행한 1D CNN 모델, 그리고 2개 온도 분류(-4℃, 3℃)에서는 IQR 및 정규화 전처리를 적용한 하이브리드 1D-2D CNN 모델이 각각 최종 선정되었다. 모델별 IG 분석 결과, 전처리 방식이나 입력 구조가 달라지더라도 반복적으로 선택된 공통 파장대가 존재하였다(Table 5).
Table 5.
Top five optimal wavelengths of the best-performing temperature classification models under each condition.
비교한 결과, 공통적으로 NIR 영역의 우세성이 확인되었다. 특히 873 - 890 nm 및 919 - 940 nm 대역은 대부분의 모델에서 지속적으로 상위 중요 파장으로 선정되었으며, 이는 저온 처리에 따른 세포 구조 변화, 수분 재분포, 조직 밀도 변화와 같은 생리·광학적 반응이 NIR 반사 특성에 안정적으로 반영됨을 시사한다(Hussain et al., 2023). 또한 일부 조건에서는 523 - 578 nm의 VIS 영역도 중요한 파장으로 도출되었는데, 이는 엽록소·카로티노이드 등 주요 색소의 흡수 특성과 연관되어 색소 변동성 및 광합성 관련 신호를 반영하는 영역으로 해석된다(Srivastava, 2021).
IG 기반 중요도 분석에서는 이러한 경향이 더욱 일관되게 나타났으며, 그중에서도 553 nm와 877 - 911 nm는 모든 모델링 조건에서 공통적으로 높은 기여도를 보인 핵심 파장대로 도출되었다. 553 nm는 엽록소와 보조색소의 주요 흡수대에 위치하여 광합성 색소 조성 변화가 반사율 변동으로 직접 연결되는 파장대이다(Gitelson et al., 2022). 반면 877 - 911 nm는 세포벽 조직, 수분 함량, 내부 구조적 변화가 민감하게 반영되는 전형적인 근적외 구조·수분 민감 대역으로, 온도 스트레스에 따른 조직 수준의 변화를 가장 안정적으로 포착하는 영역으로 판단된다(Perkov et al., 2025).
한편, IG 결과는 η2 (effect size) 분석과 비교했을 때 평가기준의 상이성으로 인해다른 양상을 보였다. η2는 파장별 반사도 분산을 기준으로 온도 변화가 반사율에 미치는 물리적 영향력을 평가하며, 이 분석에서는 400 - 420 nm, 700 - 800 nm, 그리고 900 nm 이상의 NIR 구조 민감대에서 높은 반응성이 관찰되었다. 반면 IG는 모델의 최종 분류에 기여하는 기능적 중요도를 평가하므로, 물리적 반응 범위가 넓은 파장이라 하더라도 모델 예측에 실질적 도움이 되지 않는다면 선택되지 않는다. 그 결과 IG 분석에서는 553 nm 및 877 - 911 nm와 같이 반복적이고 안정적인 신호 패턴을 제공하는 파장대가 집중적으로 도출되었다. 즉, η2는 온도 변화에 따른 스펙트럼의 물리적 민감도를, IG는 해당 밴드가 분류 정확도 향상에 기여하는 기능적 중요도를 반영한다고 해석할 수 있다.
특히 900 nm 이상의 NIR 대역은 η2와 IG 모두에서 중요성이 입증된 유일한 영역으로, 이는 세포 구조적 변화 및 수분 상태의 온도 반응이 생리적 측면과 모델적 측면에서 모두 의미 있는 신호로 작용함을 보여준다(Nur and Bachmann, 2023). 종합적으로, 553 nm와 877 - 911 nm은 복숭아 꽃의 냉해 반응을 정량적으로 반영하는 핵심 파장대로 확인되었으며, 향후 냉해 조기 진단, 스트레스 민감 파장대 탐색, 그리고 식물 기반 온도 모니터링 모델 개발에서 중요한 바이오마커 파장대로 활용될 수 있을 것으로 판단된다.
Discussion
선행연구
기존 복숭아 냉해 관련 연구는 주로 과일 단계에서 발생하는 피해를 평가하거나 화학적·생리학적 지표 기반 분석에 초점을 두었으며, 냉해 이후 과육의 갈변도, 전분·당 함량, 유기산 조성, 전도도 변화 등을 측정하는 방식으로 냉해 발생 여부를 판단하였다(Cao et al., 2023; Jia et al., 2023; Li et al., 2025; Niu et al., 2025). 이러한 접근은 냉해로 인한 품질 저하를 평가하는 데에는 유효하지만, 시료 파괴가 필수적이고 분석에 상당한 시간이 소요된다는 점에서 한계가 있다. 특히 개화기와 같이 조직 크기가 작고 변화가 미세하게 나타나는 단계에서는 조기 탐지 방법으로 활용하기 어렵다는 제약이 존재한다.
이에 비해 본 연구는 복숭아 개화시기 전 꽃 단계에서의 냉해 반응을 초분광 기반으로 직접 정량화했다는 점에서 선행연구와 명확히 차별화된다. 개화 단계는 냉해 발생에 가장 민감한 시기임에도 불구하고, 실제 연구 축적은 매우 부족한 실정이며, 대부분의 기존 연구는 과일 형성 이후 단계에서 피해를 평가해 왔다. 본 연구는 400 - 1,000 nm의 연속 스펙트럼 정보를 활용하여 온도별 반응 특성이 반사도 패턴에 어떻게 반영되는지를 분석하고, 이를 기반으로 ML 및 DL 모델을 비교하여 개화기 냉해를 고정밀로 예측한 최초의 사례 중 하나로 볼 수 있다.
또한 기존 화학적 분석 연구와 달리, 본 연구는 꽃 구조를 손상시키지 않고 비파괴·실시간 기반의 분광 지표를 활용하여 냉해 반응을 탐지할 수 있다는 장점을 지닌다. 특히 IG 기반 중요 파장 분석과 η2 기반 물리적 민감도 분석을 통해 냉해 반응에 민감한 핵심 파장대를 규명함으로써, 꽃 조직을 대상으로 한 냉해 조기 탐지 가능성을 제시하였다. 이는 화학적 구성 변화가 아닌 광학적·조직학적 반응을 반영한 정량적 지표라는 점에서 기존 연구와 근본적으로 접근 방식이 다르다.
결과적으로, 본 연구는 기존 복숭아 냉해 연구가 과일·화학분석 중심으로 제한되어 있던 범위를 확장하여, 개화기 꽃 조직에 대한 초분광 기반 AI 모델링이라는 새로운 분석 틀을 제시하였다. 이는 개화기 냉해에 대한 선제적 대응 및 재배 관리 측면에서 기존 연구의 한계를 보완하는 중요한 기여로 평가될 수 있다.
한계점
본 연구는 복숭아 꽃의 냉해 반응을 초분광 기반으로 정량화하고 AI 모델을 활용하여 온도 구분 성능을 분석하였으나, 몇 가지 한계점이 존재한다. 첫째, 사용된 데이터의 규모가 충분히 크지 않아 모델 학습 및 일반화 성능을 평가하는 데 제약이 있었다. 복숭아 꽃의 개화기 특성상 실험 반복 수와 확보 가능한 개체 수가 제한적이었고, 이는 모델이 다양한 생육 조건과 개체 간 변이를 충분히 반영하는 데 어려움을 초래했을 가능성이 있다.
둘째, 온도 조건 간 분류에서 0℃처리의 판별력이 상대적으로 낮다는 점도 중요한 제한점으로 확인되었다. 본 연구에서 0℃는 3℃와 스펙트럼 패턴이 유사하여 반복적으로 오분류되는 양상을 보였으며, 이는 경계 온도 구간에서 꽃 조직의 생리·광학적 반응 차이가 미세하여 모델이 구조적 특징을 안정적으로 학습하기 어려웠기 때문으로 해석된다. 이러한 특성은 0℃구간이 실험의 목적에 따라 분석 대상에서 제외하거나 독립 구간으로 포함할지를 결정해야 함을 시사한다. 따라서 보다 정밀한 온도 단계 구분을 위해서는 추가적인 스펙트럼 보정 또는 보조 지표 도입이 요구된다.
셋째, 본 연구는 단일 촬영 환경과 제한된 한 품종을 기반으로 수행되었기 때문에, 실제 재배환경의 다양한 온도 변동, 빛 조건, 품종 다양성 등을 반영하는 데에는 한계가 있다. 복숭아 꽃의 냉해 반응은 품종별로 차이가 존재할 수 있으므로, 향후 다품종 비교 실험과 다양한 환경 조건에서의 반복 검증이 요구된다.
마지막으로, 본 연구에서 구축한 모델은 초분광 영상만을 기반으로 개발되었기 때문에, 구조적·생리적 변화를 종합적으로 반영하기에는 부족할 수 있다. 냉해는 조직 내부 수분 변화, 세포막 손상 등 비가시적 요인이 함께 작용하므로, 이를 보완하기 위해 열영상, RGB, 전기적·기계적 센서와의 멀티모달 통합이 향후 연구에서 고려되어야 한다 판단된다.
향후연구
앞서 제시한 한계점을 보완하기 위해서는 몇 가지 후속 연구가 필요하다. 첫째, 데이터 규모 확장은 필수적이다. 개체 수와 반복 실험 수를 증가시키고, 여러 생육 단계와 다양한 개체 변이를 반영한 대규모 데이터셋을 구축함으로써 모델의 일반화 성능을 강화할 수 있을 것이다. 특히 개화기 샘플의 확보가 제한적이었던 점을 고려할 때, 온실·재배지 등 다양한 환경에서 장기간 반복 촬영을 수행하는 체계적 데이터 수집이 요구된다.
둘째, 0℃와 3℃와 같이 스펙트럼 분리도가 낮은 경계 온도 구간의 판별력을 높이기 위해 보다 정교한 전처리 기법과 추가적인 생리·광학 지표의 도입이 필요하다. 파장 해상도 보완, 비선형적 특징 변환, 대역 확장 등을 포함한 고급 분광 처리 기법이 해당 문제를 완화할 수 있으며, 비가시적 조직 변화를 반영하는 보조 센서를 활용한 멀티모달 융합 또한 유효한 접근이 될 것이다.
셋째, 본 연구에서는 단일 촬영 환경과 제한된 품종을 대상으로 실험을 수행하였으므로, 향후에는 다양한 광원 조건, 배경 환경, 개화 단계, 그리고 복수 품종을 포함하는 확장 연구가 필요하다. 품종 간 냉해 민감도 차이를 반영한 대조 실험은 모델의 적용 가능 범위를 넓히고, 품종 맞춤형 냉해 진단 시스템 개발로 이어질 수 있다.
넷째, 초분광 영상 기반의 단일 입력 구조에서 벗어나, RGB·열영상·근적외선·LiDAR 기반 구조 정보 등을 결합하는 멀티모달 모델링이 향후 유망한 방향으로 제시된다. 이러한 통합 모델은 꽃 조직의 색소 변화, 수분 분포, 구조적 손상 등을 함께 반영할 수 있어 예측 안정성과 조기 탐지 능력을 더욱 향상시킬 것으로 기대된다.
마지막으로, 현장에서 실시간 진단에 활용할 수 있는 경량화 모델과 이동형 촬영 시스템 개발도 중요한 연구 주제이다. 경량 초분광 센서 또는 RGB + 근적외선 기반 다중 스펙트럴 카메라를 활용하면 재배지에서 빠른 냉해 진단이 가능하며, 이는 농가 의사결정 지원 및 냉해 피해 예방 전략 수립에 직접적인 기여를 할 수 있을 것이라 판단된다.
Conclusion
본 연구에서는 복숭아 꽃에서 발생하는 냉해 반응을 조기에 판별하기 위해 초분광 영상 기반의 분석 체계와 AI 온도 분류 모델을 구축하였다. 400 - 1,000 nm 범위의 초분광 영상에서 ROI 기반 스펙트럼을 추출하고 표준화된 전처리 절차를 적용하여 분석용 입력을 구성하였으며, 이를 통해 여러 온도 조건별을 구분하는 분류 모델을 개발하였다. 그 결과, IQR 전처리와 CNN 조합이 모든 온도 클래스에서 가장 안정적이고 우수한 성능을 보였으며, 이는 꽃 조직의 미세한 온도 반응 차이를 효과적으로 학습한 결과로 판단된다.
평균 스펙트럼 분석에서는 3℃ 이하의 저온에서 반사도가 전반적으로 감소하는 경향이 관찰되었으며, 이는 저온 스트레스에 따른 세포 구조 약화 및 수분 감소와 관련된 광학적 변화로 해석된다. η2 기반 민감도 분석에서는 전체 파장대에서 0.10 - 0.30 범위를 나타냈고, 특히 400 - 420 nm, 700 - 800 nm, 900 nm 이상에서 민감도가 높게 나타나 온도 반응에 중요한 스펙트럼 영역임을 확인하였다.
모델 기반 중요도 분석에서도 520 - 525 nm, 575 - 580 nm, 675 - 680 nm, 785 - 790 nm, 870 - 880 nm, 890 - 920 nm, 그리고 약 940 nm 인근 파장대가 반복적으로 중요한 특징으로 도출되었다. 특히 675 - 680 nm, 785 - 790 nm, 870 - 880 nm 및 890 - 920 nm은 냉해 반응과 생리적 변화가 직접적으로 반영되는 영역으로 확인되었다. 이를 종합하면, 400 - 420 nm, 675 - 680 nm, 785 - 790 nm, 870 - 900 nm 영역은 복숭아 꽃의 저온 반응 감지에 핵심적인 스펙트럼 범위로 제안될 수 있다.
종합적으로, 본 연구는 기존 과일 단계의 냉해 평가 또는 화학적 분석에 머물렀던 선행연구의 한계를 넘어, 개화기 복숭아 꽃에서 발생하는 저온 반응을 비파괴적으로 조기에 탐지할 수 있는 새로운 분석 프레임워크를 제시하였다. 본 연구의 결과는 복숭아 냉해 조기경보 시스템 구축, 저온 내성 품종 개발, 재배지 의사결정 지원 등 다양한 응용 가능성을 갖는 기반 자료로 활용될 수 있다.
