

Introduction
Materials and Methods
연구대상지 및 연구수종
실험 설계
토양 분석
생장 및 광합성 측정
잎 양분특성
통계분석
Results and Discussion
시비수준에 따른 수종별 토양 특성
시비수준에 따른 수종별 생장변화
Conclusion
Introduction
기후변화로 인한 기온 상승으로 우리나라의 산림식생대는 점차 북상하고 있어, 남쪽 지역의 난대림 지대가 북쪽으로 확대될 것으로 예측되고 있다. 실제로 1990년대 중반 이후 20년간 침엽수림 면적은 약 25% 감소한 것으로 나타났으며, 2055년까지 국내 침엽수림이 전체 임상 중 38.1%에서 28.5%까지 감소할 것이라고 예측하고 있다(Kim et al., 2022). 결과적으로, 우점하고 있던 소나무 등 침엽수종과 한대성 수종의 면적이 줄고 난대수종의 면적이 증가하게 될 전망이다. 이에 따라 우리나라에서는 남부 지방을 중심으로 난대수종이 조림되고 있으며, 점차 중부지역으로 확대될 것으로 예상된다(KFS, 2009). 난대수종의 생육 범위가 확대됨에 따라 건전한 임분을 조성하는 것은 필수적인 과제이다. 이를 위해서는 조림지에서 묘목의 적응성과 초기 생장률을 높이고, 양묘단계에서 적절한 양분관리를 통해 건전한 묘목 생산이 뒷받침되어야 한다(Carlson, 1981; Imo and Timmer, 1999; Quoreshi and Timmer, 2000).
시비를 통해 토양으로 유입된 양분은 이온화되어 토양입자 및 유기물에 부착되거나 토양미생물 또는 식물체에 이용된다. 질소 비료의 이온화 속도는 높고 인은 낮은 특성이 있으며, 토양 조건에 따라 이온화된 양분의 이동성은 달라진다(Brady and Weil, 2002). 이러한 원소의 이동성 차이는 식물의 영양분 가용성에 영향을 미친다(Havlin et al., 2016). 따라서 적절한 시비 사용은 토양의 양분 보충과 임목의 생장효과를 기대할 수 있다(Son et al., 2020). NPK 시비에 따른 가시나무류 생장특성에 관한 연구에서는 간장과 근원경이 수종에 상관없이 시비수준이 높아질수록 생장이 증가한다고 보고되었다(Kim et al., 2014). 그러나 Park 등(2019)의 시비처리에 따른 편백나무의 생장 연구에서는 시비 강도 처리구의 묘목의 생장량이 약도와 중도 처리구보다 낮게 나타난다고 보고되었다. 또한 시비수준이 높은 경우에는 묘목의 품질을 떨어뜨릴 수 있어, 과도한 시비는 묘목의 생장에 악영향을 끼칠 수 있다(Cha et al., 2019). 또한 포지에서 수종과 토양환경을 고려하지 않은 화학비료의 사용은 토양의 질을 저감 시키고, 발아율과 묘목 생장을 저하시키며, 포지 주변의 토양과 지하수의 오염을 유발할 수 있다(Broschat, 1995; Andersen and Hansen, 2000). 따라서 이를 방지하고, 건전한 묘목의 생산을 위해서는 수종과 포지 토양 조건을 고려한 비료 종류와 시비량의 기준 정립이 요구된다.
식물의 생장에 있어 질소와 인은 중요한 무기양분이다(Zhang et al., 2017). 그 중 질소는 가장 많은 양이 식물체 내 함유되어 있어 가장 중요한 원소이며, 식물의 단백질, 핵산, 엽록소 등을 구성하고 광합성과 호흡 등의 대사작용을 왕성하게 한다. 질소가 결핍되면, 잎의 황화현상이 시작되고, 지상부의 생장이 저조하여 영양생장이 지연된다(Poole, 1978). 또한 질소는 일반적인 풍화작용으로 생성되지 않기 때문에 다른 원소에 비해 비교적 결핍이 쉽다는 특징이 있다. 따라서 양묘 측면에서 질소 시비는 수목의 생장에 큰 영향을 미치기 때문에 필수적이라고 할 수 있다(Binkley, 1986; Kozlowski and Pallardy, 1997).
인은 에너지를 생산하고 전달하는 과정에 직접 관여하며, 광합성과 호흡작용에서 여러 가지 대사를 주도한다(Lee, 2021). 특히 뿌리 발달에 관여하여 전반적인 생장에 영향을 미치는데, specific root length (SRL)와 specific root area (SRA)를 증가시켜 양분흡수 능력이 향상되어 건강한 묘목 발달을 촉진한다(Jin et al., 2024). 인이 결핍되면 묘목의 초기생장단계에서 신초의 생장이 불량해지고 뿌리발달과 병해에 대한 저항력이 저하되는 것으로 알려져 있다(Ma, 1979). 임목의 인 요구도는 질소에 비하여 상대적으로 훨씬 낮지만, 토양 내 인의 가용성은 매우 낮기 때문에, 1년 요구량 대비 유입량의 비율은 훨씬 높다(Son et al., 2020). 따라서 이러한 양분의 특성과 수종 및 토양 환경 조건에 맞는 시비를 적용하는 것은 중요하다.
현재 우리나라에서는 묘목의 생장과 안정적인 활착을 위해 시비수준을 달리하여 양묘하고 있으며, 그 효과에 관한 연구는 많이 보고되고 있다(Son et al., 1998; Shin et al., 1999; Park et al., 2010). 하지만 우리나라의 난대수종 조림범위가 북상하고 있음에도, 묘목 생산과정에 도입되는 시비처리 수준에 관한 연구는 미흡한 상황으로, 이에 대한 지속적인 연구가 요구된다.
본 연구에서는 산림청 권장조림수종 중 난대수종인 편백, 후박나무, 참가시나무의 적정 시비 수준을 구명하기 위해 질소와 인 비료 함량을 다르게 한 16가지 수준의 시비처리를 실시하였다. 처리 별 효과를 알아보기 위해 수종별로 묘목의 생장특성(근원경, 묘고), 엽면적비(specific leaf area, SLA), 광합성량, 잎의 양분특성, 토양양분 특성을 조사하였다. 이를 통해 편백, 후박나무, 참가시나무의 묘목 생산과정에서 최적의 시비수준을 정립시키고, 더욱 효율적인 우량 묘목생산이 이루어질 수 있을 것이라고 기대된다.
Materials and Methods
연구대상지 및 연구수종
본 연구는 전라남도 보성에 위치한 서부지방산림청 보성양묘사업소(36°44′55.83″N, 127°11′00.86″E)에서 편백, 후박나무, 참가시나무 1-0 묘목을 대상으로 실시하였다. 편백 묘목은 전남 보성의 보성양묘사업소에서, 후박나무 묘목은 광양에서, 참가시나무 묘목은 제주 난대아열대산림연구소에서 생산되었다. 수종별 초기 근원경과 묘고는 Table 1과 같다. 이 지역의 평균기온은 14.0℃, 연평균강수량은 1,410 mm이다(KMA, 2022).
Table 1.
Initial root collar diameter and height of three species.
| Species | Root collar diameter (mm) | Height (cm) |
| Chamaecyparis obtusa | 1.37 ± 0.26 | 8.62 ± 1.22 |
| Machilus thunbergii | 1.90 ± 0.27 | 9.35 ± 1.40 |
| Quercus salicina | 2.66 ± 0.66 | 8.33 ± 2.43 |
실험 설계
양묘장 내 너비 1 m, 길이 20 m 포지 15개를 동서 방향으로 배치하고, 상 위에 1 m × 1 m의 처리구를 16개씩 만들었다. 조사구 사이에는 50 cm 거리를 두어 작업로와 배수로로 이용하였다. 각 포지 당 16개의 처리를 완전임의배치하였으며, 전체 포지를 3등분하여 각 수종별로 5개의 포지에 각각 편백, 후박나무, 참가시나무를 식재하였다. 전체 실험은 16처리, 5반복, 3수종으로 총 240개의 시험구에서 진행되었다. 시비처리는 질소비료로 요소, 인 비료로 용과린을 사용하였으며, 산림청 종묘사업실시요령 표준시비량인 요소 30 g·m-2 (N 13.8 g·m-2), 용과린 70 g·m-2 (P 6.1 g·m-2)을 1배량의 기준으로 설정하였다. 질소 4수준(0, 1, 2, 3배량), 인 4수준(0, 1, 2, 3배량)을 조합하여 16개의 처리를 3가지 수종에 5반복씩 적용하였다(Table 2).
3월 말부터 4월 초 사이에 생육상태가 비교적 유사한 묘목을 선정하여, 1 m × 1 m 처리구 240개에 하루에 걸쳐 심도록 하였다. 식재 전, 편백과 참가시나무는 단근작업을 수행하였으며, 묘목의 뿌리 건조를 막고 활착률을 높이기 위해 뿌리를 황톳물에 담근 후 식재하였다. 각 수종별로 산림청 양묘사업기준에 따라 한 처리구에 편백은 식재거리 13 cm, 식재본수 30본, 후박나무는 식재거리 11 cm, 식재본수 20본, 참가시나무는 식재거리 13 cm, 식재본수 12본으로 설정하였다.
식재 후, 추위에 취약한 후박나무는 볏짚을 이용하여 5 - 7 cm 두께의 멀칭을 수행하였고, 바람에 날리지 않도록 새끼줄을 통해 고정하였다. 스프링쿨러를 이용하여 관수하였고, 잡초 제거는 봄철은 3주에 한번 그리고 여름 동안은 2주에 한번 꼴로 시행하였다. 또한 차광막을 이용하여 바람막이를 설치하였다. 시비는 식재 4주 후, 묘목사이에 흩어뿌림 방식으로 처리하였다.
Table 2.
Amount of nitrogen and phosphorus per 1 m2 area of fertilizer applied according to 16 treatments.
토양 분석
시비 처리 전, 토양의 물리·화학적 특성을 분석하기 위해, 수종별 포지의 대각선에 따라 일정한 간격으로 3개의 지점에서 0 - 10 cm 깊이의 토양시료를 채취하여 분석하였다. 시비 처리 후, 각 처리구에서 무작위로 선택한 두 지점의 토양 표층의 유기물을 제거한 후 0 - 10 cm 토양 깊이에서 토양시료를 500 g씩 채취하고, 처리별로 2반복 분석하였다.
시비처리 전과 후는 동일한 분석방법을 사용하였으며, 토양 분석을 위해 토양시료를 분석 전까지 4℃에서 보관하였다. 토성은 30℃에서 hydrometer법을 사용하였으며, 유기물 함량은 Tyurin법으로 측정하였다. 토양산도 측정은 pH meter법을 이용하였으며, 전질소는 Micro-Kjeldahl법, 유효인산(P2O5)은 Lancaster법을 이용하였다. 치환성 양이온 K+, Ca2+, Mg2+은 1 N의 NH4OAc을 이용해 측정한 뒤 atomic absorption spectrometer (AA280FS, Agilent Technologies, USA)을 사용해 측정하였다. 양이온교환용량(cation exchange capacity, CEC)는 1 N의 NH4OAc와 CH3COOH용액으로 양이온을 추출한 후 이용하여 Brown법으로 측정하였다.
생장 및 광합성 측정
시비처리에 따른 생장반응을 측정하기 위해 시비처리 전과 처리 5개월 후에 세 수종을 대상으로 근원경과 묘고를 측정하였다. 근원경은 전자식 캘리퍼스(Absolute Digimatic Caliper-500, Mitutoyo, Japan)를 이용하여 지상으로부터 1 cm 높이에서 측정하였으며, 측정한 자리에 흰색 페인트로 표시하여 처리 전과 처리 후에 동일한 위치를 측정할 수 있도록 하였다. 묘고는 정단까지 접이식 자를 이용하여 측정하였다. 측정된 데이터를 가지고 시비처리 후의 생장량을 계산하고, 아래의 상대 생장률 계산식(식(1), (2))을 사용해서 근원경과 묘고의 상대생장률을 구하였다(Hunt, 1978).
: relative height growth rate
: relative root collar diameter growth rate
: height
: root collar diameter
: time
엽면적비(SLA) 측정을 위한 엽면적은 개체 당 정단 부위부터 3 - 4번째 잎을 2 - 4개씩 채취하여 엽면적 측정기(LI-3100, LI-COR Inc., USA)로 측정하였다. 이후, 85℃에서 7일 건조 후 건중량을 측정하였다. SLA는 엽면적을 건중량으로 나누어 아래의 식(3)으로 계산하였다(Hunt, 1978).
시비 수준이 다른 조건에서 생장한 묘목들의 광합성 반응 측정은 식재 후 약 5개월이 지난 시점에 실시하였다. 측정 대상은 처리별 생장 속도가 비슷하고 평균적인 생육 상태를 유지하고 있는 묘목의 건전한 잎으로 선정하였다. 광합성 측정은 휴대용 광합성 측정기(LI-6400, LI-COR Inc., USA)를 사용하여 수행하였다. 각 처리 내 변동성을 최소화하기 위해 동일한 시간대에 측정을 진행하였으며, 수종별로 무작위로 선정된 5개체를 대상으로 광포화점(1,000 µmol·m-2·s-1)에서 광합성 반응을 측정하였다. 잎 챔버에 유입되는 CO2 농도는 400 µmol·mol-1·air로 유지하였고, 챔버 내 온도는 25℃, 상대 습도는 40 - 50%로 조절하였다.
잎 양분특성
잎의 양분 분석을 위해 잎을 채취하였으며, 가장자리에 위치한 묘목의 경우 가장자리 효과를 제거하기 위해 내부 방향으로 뻗은 잎을 선정하였다. 건조한 잎을 곱게 간 후, H2SO4와 ClO4 혼합용액을 이용한 block digestor (BD-46, Lachat Ins., USA) 방법을 사용하여 유기물을 분해하였다. 전처리를 마친 시료는 automated ion analyzer (Quik Chem AE, Lachat Ins., USA)를 이용해 N, P 농도를 측정하였으며, atomic absorption spectrometer를 이용해 K, Ca, Mg 농도를 측정하였다.
통계분석
시비처리가 편백, 후박나무, 참가시나무 묘목의 생장과 양분 흡수에 미치는 영향을 분석하기 위해 R (version 4.1.1, Austria)을 이용하여 통계분석을 실시하였다. 각 수준별 시비처리가 묘목 생장량, 광합성과 잎 양분에 미치는 영향을 분석하기 위해 이원분산분석(two-way ANOVA)을 실시하였으며, 각 처리 간의 유의성은 다중 범위 검정(Duncan’s multiple range test)을 유의수준 5%에서 분석하였다(p < 0.05).
Results and Discussion
시비수준에 따른 수종별 토양 특성
시비 처리 전, 토양의 물리·화학적 특성은 Table 3과 같다. 세 수종의 포지 모두 각 항목의 값이 비슷한 수치로 조사되었다.
Table 3.
Soil properties before treatments.
| Properties | Depth (cm) | Chamaecyparis obtusa | Machilus thunbergii | Quercus salicina |
| Sand (%) | 0 - 10 | 61.71 (0.39)z | 59.10 (0.26) | 57.39 (0.75) |
| 10 - 20 | 60.71 (0.51) | 58.64 (1.07) | 56.50 (0.41) | |
| Silt (%) | 0 - 10 | 28.55 (0.49) | 30.62 (0.26) | 30.41 (0.85) |
| 10 - 20 | 29.12 (0.40) | 30.76 (0.97) | 30.65 (0.55) | |
| Clay (%) | 0 - 10 | 9.75 (0.11) | 10.28 (0.00) | 12.21 (0.49) |
| 10 - 20 | 10.17 (0.11) | 10.60 (0.18) | 12.85 (0.56) | |
| pH (-Log [H+]) | 0 - 10 | 5.17 (0.02) | 5.24 (0.07) | 5.36 (0.02) |
| 10 - 20 | 5.15 (0.01) | 5.26 (0.02) | 5.32 (0.02) | |
| Organic matter (%) | 0 - 10 | 1.13 (0.03) | 1.12 (0.02) | 1.41 (0.10) |
| 10 - 20 | 1.18 (0.03) | 1.23 (0.02) | 1.45 (0.16) | |
| Total N (g·kg-1) | 0 - 10 | 1.30 (0.12) | 1.03 (0.03) | 1.10 (0.06) |
| 10 - 20 | 1.07 (0.12) | 1.10 (0.15) | 1.07 (0.09) | |
| Available Py (mg·kg-1) | 0 - 10 | 150.60 (8.75) | 151.80 (5.32) | 170.54 (15.89) |
| 10 - 20 | 153.77 (7.69) | 153.68 (5.85) | 177.93 (16.40) | |
| Exchangeable K+ (cmolc·kg-1) | 0 - 10 | 0.55 (0.02) | 0.50 (0.03) | 0.55 (0.04) |
| 10 - 20 | 0.53 (0.03) | 0.49 (0.02) | 0.52 (0.05) | |
| Exchangeable Ca2+ (cmolc·kg-1) | 0 - 10 | 1.81 (0.09) | 2.15 (0.10) | 2.56 (0.10) |
| 10 - 20 | 1.73 (0.09) | 2.16 (0.13) | 2.59 (0.14) | |
| Exchangeable Mg+ (cmolc·kg-1) | 0 - 10 | 0.70 (0.04) | 0.78 (0.02) | 0.75 (0.03) |
| 10 - 20 | 0.67 (0.05) | 0.76 (0.01) | 0.77 (0.02) | |
| CEC (cmolc·kg-1) | 0 - 10 | 8.58 (0.66) | 7.99 (0.90) | 9.42 (0.55) |
| 10 - 20 | 7.92 (0.35) | 7.26 (0.34) | 9.86 (0.60) |
시비 처리 후의 토양 특성은 수종 보다는 비료의 종류와 시비 수준에 따른 변화를 보였다(Tables 4, 5, 6). 이전 연구들이 화학비료의 시비가 토양 산성화를 촉진시킨다고 보고한 것처럼(Um et al., 2019), 대체로 시비량이 증가할수록 토양 pH는 감소하는 경향이 나타난다. 특히 질소 시비가 증가할수록 그 차이가 크게 나타나는데, 이는 질소 시비가 토양 내 수소이온을 증가시키기 때문으로 보인다(Barak et al., 1997). 총 질소량은 시비처리 후 감소했으나, 질소비료의 시비량이 늘어날수록 증가하는 모습을 보이며 그 차이가 크지 않았다. 이러한 결과는 타 연구와 같이 시비처리에 의한 질소함량은 크게 변하지 않으며, Miller (1981)의 연구에 따르면, 시비된 질소가 식물에 생장에 이용되거나, 질산의 형태로 탈질화 되기 때문에 처리 전 보다 그 양이 적어 질소 수준에 도움을 주지 못한 것으로 판단된다. 질소 시비량의 증가로 인한 수소이온의 증가는 치환성 양이온의 용탈을 야기하여 치환성 칼슘과 마그네슘이 감소하는 현상을 초래한 것으로 보인다(Kwon et al., 1998). 또한 인 시비량이 증가할수록 치환성 칼슘과 마그네슘이 증가하는데, 이는 인 비료로 사용한 용과린의 석회 성분으로 토양 산성화가 중화되어 나타나는 현상으로 사료된다.
가용성 인의 경우, 질소 시비에 따른 유의한 차이를 보이지 않았지만, 인 시비량이 증가할수록 증가했다. 이는 토양수에 용해되어 식물이 흡수할 수 있는 수용성 인의 농도가 증가하기 때문으로 사료된다(Bindraban et al., 2020).
Table 4.
Soil properties after treatments about Chamaecyparis obtusa.
| Properties |
Nitrogen (g·m-2) | Phosphorus (g·m-2) | |||
| 0 | 6.1 | 12.2 | 18.3 | ||
|
pH (-Log [H+]) | 0 | 5.22 (0.03)z | 5.15 (0.09) | 5.00 (0.07) | 4.88 (0.02) |
| 13.8 | 4.96 (0.09) | 4.76 (0.03) | 4.79 (0.06) | 4.68 (0.04) | |
| 27.6 | 4.75 (0.03) | 4.64 (0.11) | 4.46 (0.20) | 4.42 (0.01) | |
| 41.4 | 4.26 (0.16) | 4.53 (0.17) | 4.29 (0.06) | 4.36 (0.13) | |
|
Total N (g·kg-1) | 0 | 0.82 (0.00) | 0.71 (0.00) | 0.83 (0.01) | 0.76 (0.01) |
| 13.8 | 0.81 (0.06) | 0.81 (0.01) | 0.81 (0.05) | 0.79 (0.03) | |
| 27.6 | 0.79 (0.01) | 0.85 (0.01) | 0.86 (0.03) | 0.97 (0.04) | |
| 41.4 | 0.83 (0.01) | 0.82 (0.10) | 1.08 (0.09) | 1.05 (0.06) | |
|
Available Py (mg·kg-1) | 0 | 123.90 (4.23) | 173.49 (13.54) | 239.62 (34.93) | 274.26 (17.22) |
| 13.8 | 126.02 (16.55) | 171.17 (16.30) | 200.69 (39.66) | 243.47 (20.52) | |
| 27.6 | 121.28 (10.73) | 166.75 (0.90) | 192.51 (2.40) | 317.86 (21.06) | |
| 41.4 | 126.25 (6.66) | 199.78 (35.58) | 210.84 (41.09) | 270.54 (28.17) | |
|
Exchangeable K+ (cmolc·kg-1) | 0 | 0.60 (0.00) | 0.63 (0.03) | 0.65 (0.03) | 0.66 (0.05) |
| 13.8 | 0.57 (0.11) | 0.57 (0.08) | 0.58 (0.05) | 0.51 (0.00) | |
| 27.6 | 0.59 (0.05) | 0.56 (0.00) | 0.51 (0.03) | 0.63 (0.04) | |
| 41.4 | 0.47 (0.06) | 0.53 (0.06) | 0.66 (0.05) | 0.63 (0.04) | |
|
Exchangeable Ca2+ (cmolc·kg-1) | 0 | 1.46 (0.07) | 1.38 (0.13) | 1.46 (0.16) | 1.39 (0.06) |
| 13.8 | 1.04 (0.23) | 0.91 (0.05) | 1.22 (0.01) | 1.10 (0.11) | |
| 27.6 | 0.89 (0.07) | 0.83 (0.06) | 0.95 (0.01) | 1.06 (0.19) | |
| 41.4 | 0.77 (0.05) | 0.87 (0.10) | 1.31 (0.06) | 0.93 (0.04) | |
|
Exchangeable Mg+ (cmolc·kg-1) | 0 | 0.59 (0.03) | 0.60 (0.04) | 0.63 (0.06) | 0.63 (0.03) |
| 13.8 | 0.42 (0.08) | 0.43 (0.01) | 0.52 (0.01) | 0.50 (0.00) | |
| 27.6 | 0.38 (0.04) | 0.40 (0.05) | 0.52 (0.01) | 0.62 (0.05) | |
| 41.4 | 0.38 (0.05) | 0.42 (0.03) | 0.71 (0.08) | 0.58 (0.06) | |
|
EC (dS·m-1) | 0 | 0.39 (0.01) | 0.37 (0.00) | 0.42 (0.00) | 0.41 (0.02) |
| 13.8 | 0.30 (0.02) | 0.36 (0.02) | 0.43 (0.04) | 0.49 (0.00) | |
| 27.6 | 0.42 (0.07) | 0.45 (0.10) | 0.77 (0.19) | 1.12 (0.08) | |
| 41.4 | 0.91 (0.37) | 0.53 (0.08) | 1.86 (0.58) | 1.63 (0.25) | |
Table 5.
Soil properties after treatments about Machilus thunbergii.
| Properties |
Nitrogen (g·m-2) | Phosphorus (g·m-2) | |||
| 0 | 6.1 | 12.2 | 18.3 | ||
|
pH (-Log [H+]) | 0 | 5.45 (0.01)z | 5.22 (0.10) | 5.13 (0.05) | 4.92 (0.09) |
| 13.8 | 5.26 (0.01) | 5.11 (0.14) | 4.93 (0.12) | 4.52 (0.04) | |
| 27.6 | 4.90 (0.10) | 4.83 (0.05) | 4.63 (0.04) | 4.40 (0.04) | |
| 41.4 | 4.90 (0.10) | 4.61 (0.09) | 4.34 (0.11) | 5.20 (0.09) | |
|
Total N (g·kg-1) | 0 | 0.80 (0.06) | 0.80 (0.07) | 0.78 (0.00) | 0.76 (0.03) |
| 13.8 | 0.82 (0.01) | 0.79 (0.01) | 0.83 (0.02) | 0.94 (0.00) | |
| 27.6 | 0.88 (0.01) | 0.85 (0.01) | 0.88 (0.02) | 1.04 (0.02) | |
| 41.4 | 0.83 (0.02) | 0.90 (0.02) | 1.02 (0.10) | 0.92 (0.04) | |
|
Available Py (mg·kg-1) | 0 | 130.21 (10.17) | 176.83 (3.36) | 234.79 (27.85) | 315.93 (39.19) |
| 13.8 | 127.37 (5.05) | 196.76 (30.03) | 243.05 (17.11) | 313.97 (9.08) | |
| 27.6 | 132.88 (8.73) | 189.97 (1.22) | 239.33 (17.06) | 284.13 (11.11) | |
| 41.4 | 124.29 (10.60) | 186.13 (11.51) | 188.82 (4.34) | 185.29 (16.88) | |
|
Exchangeable K+ (cmolc·kg-1) | 0 | 0.95 (0.01) | 0.92 (0.01) | 0.71 (0.07) | 0.69 (0.05) |
| 13.8 | 0.89 (0.07) | 0.81 (0.07) | 0.82 (0.01) | 0.70 (0.00) | |
| 27.6 | 0.78 (0.06) | 0.71 (0.07) | 0.66 (0.08) | 0.67 (0.04) | |
| 41.4 | 0.76 (0.11) | 0.77 (0.10) | 0.60 (0.08) | 0.96 (0.04) | |
|
Exchangeable Ca2+ (cmolc·kg-1) | 0 | 1.90 (0.16) | 1.99 (0.18) | 1.95 (0.30) | 1.49 (0.18) |
| 13.8 | 1.35 (0.22) | 1.58 (0.03) | 1.46 (0.35) | 1.67 (0.08) | |
| 27.6 | 1.36 (0.04) | 1.44 (0.11) | 1.60 (0.13) | 1.80 (0.05) | |
| 41.4 | 1.03 (0.01) | 1.49 (0.03) | 1.85 (0.28) | 2.21 (0.00) | |
|
Exchangeable Mg+ (cmolc·kg-1) | 0 | 0.59 (0.05) | 0.65 (0.04) | 0.66 (0.10) | 0.52 (0.07) |
| 13.8 | 0.42 (0.04) | 0.49 (0.03) | 0.50 (0.09) | 0.67 (0.02) | |
| 27.6 | 0.40 (0.02) | 0.45 (0.02) | 0.56 (0.02) | 0.71 (0.01) | |
| 41.4 | 0.34 (0.01) | 0.50 (0.02) | 0.71 (0.15) | 0.67 (0.03) | |
|
EC (dS·m-1) | 0 | 0.34 (0.03) | 0.37 (0.01) | 0.42 (0.05) | 0.53 (0.09) |
| 13.8 | 0.33 (0.03) | 0.35 (0.04) | 0.45 (0.08) | 0.44 (0.04) | |
| 27.6 | 0.50 (0.07) | 0.56 (0.06) | 0.79 (0.05) | 1.11 (0.06) | |
| 41.4 | 0.41 (0.05) | 0.80 (0.11) | 1.65 (0.77) | 1.44 (0.27) | |
Table 6.
Soil properties after treatments about Quercus salicina.
| Properties |
Nitrogen (g·m-2) | Phosphorus (g·m-2) | |||
| 0 | 6.1 | 12.2 | 18.3 | ||
|
pH (-Log [H+]) | 0 | 5.20 (0.09)z | 5.18 (0.17) | 4.77 (0.03) | 5.08 (0.08) |
| 13.8 | 5.35 (0.02) | 5.20 (0.00) | 4.78 (0.20) | 5.03 (0.09) | |
| 27.6 | 4.71 (0.09) | 4.72 (0.24) | 4.76 (0.38) | 4.83 (0.25) | |
| 41.4 | 4.48 (0.02) | 4.42 (0.11) | 4.52 (0.07) | 4.70 (0.40) | |
|
Total N (g·kg-1) | 0 | 0.92 (0.04) | 1.00 (0.17) | 1.00 (0.09) | 0.97 (0.06) |
| 13.8 | 0.87 (0.12) | 0.90 (0.08) | 0.95 (0.06) | 0.94 (0.01) | |
| 27.6 | 0.99 (0.09) | 0.97 (0.03) | 0.99 (0.03) | 0.89 (0.06) | |
| 41.4 | 0.90 (0.10) | 1.05 (0.01) | 1.02 (0.03) | 0.96 (0.07) | |
|
Available Py (mg·kg-1) | 0 | 185.29 (16.88) | 227.55 (13.95) | 290.72 (15.87) | 261.46 (11.78) |
| 13.8 | 133.21 (10.00) | 204.49 (8.83) | 273.39 (38.21) | 305.09 (29.55) | |
| 27.6 | 147.05 (19.09) | 221.38 (15.37) | 238.29 (55.32) | 198.41 (3.72) | |
| 41.4 | 149.10 (23.91) | 182.70 (33.08) | 201.78 (12.85) | 236.77 (49.18) | |
|
Exchangeable K+ (cmolc·kg-1) | 0 | 0.96 (0.06) | 1.14 (0.14) | 0.93 (0.12) | 0.83 (0.01) |
| 13.8 | 1.09 (0.11) | 0.89 (0.06) | 0.77 (0.08) | 0.84 (0.09) | |
| 27.6 | 1.15 (0.09) | 0.89 (0.11) | 0.86 (0.02) | 0.75 (0.03) | |
| 41.4 | 0.90 (0.02) | 0.86 (0.02) | 0.78 (0.03) | 0.81 (0.09) | |
|
Exchangeable Ca2+ (cmolc·kg-1) | 0 | 2.21 (0.00) | 2.20 (0.29) | 2.16 (0.10) | 2.01 (0.30) |
| 13.8 | 1.69 (0.19) | 1.82 (0.09) | 1.80 (0.24) | 2.06 (0.23) | |
| 27.6 | 1.57 (0.03) | 1.78 (0.01) | 2.02 (0.20) | 1.81 (0.10) | |
| 41.4 | 1.68 (0.07) | 1.93 (0.18) | 2.17 (0.06) | 2.08 (0.30) | |
|
Exchangeable Mg+ (cmolc·kg-1) | 0 | 0.67 (0.03) | 0.68 (0.07) | 0.72 (0.00) | 0.64 (0.08) |
| 13.8 | 0.44 (0.02) | 0.51 (0.00) | 0.59 (0.11) | 0.63 (0.03) | |
| 27.6 | 0.44 (0.01) | 0.55 (0.09) | 0.65 (0.14) | 0.55 (0.06) | |
| 41.4 | 0.52 (0.00) | 0.62 (0.07) | 0.78 (0.01) | 0.69 (0.18) | |
|
EC (dS·m-1) | 0 | 0.54 (0.04) | 0.51 (0.14) | 0.95 (0.08) | 0.50 (0.03) |
| 13.8 | 0.35 (0.04) | 0.37 (0.01) | 0.73 (0.32) | 0.43 (0.03) | |
| 27.6 | 0.91 (0.03) | 1.13 (0.6) | 1.10 (0.57) | 0.61 (0.21) | |
| 41.4 | 1.22 (0.19) | 1.49 (0.28) | 1.48 (0.19) | 1.45 (0.98) | |
시비수준에 따른 수종별 생장변화
근원경 및 묘고 생장량과 상대 생장률
편백에서 근원경 생장은 질소와 인의 복합처리에 따른 유의한 차이를 보였지만(p < 0.001), 인 시비는 유의한 영향을 주지 못했다. 후박나무와 참가시나무의 근원경 생장은 복합처리와 인 시비 모두 유의하지 않았다. 질소 시비는 세 수종 모두 유의한 영향을 주었다. 질소 단일 처리에서의 근원경 생장량은 편백이 질소 2배(N2P0)에서 가장 높았으며, 대조구(N0P0)보다 약 1.3배 높았다. 후박나무는 질소 1배(N1P0)에서 가장 높았으며, 대조구보다 약 1.4배 높았다. 참가시나무는 질소 3배에서(N3P0) 가장 높았으며, 대조구보다 약 1.1배 높았다(Fig. 1).
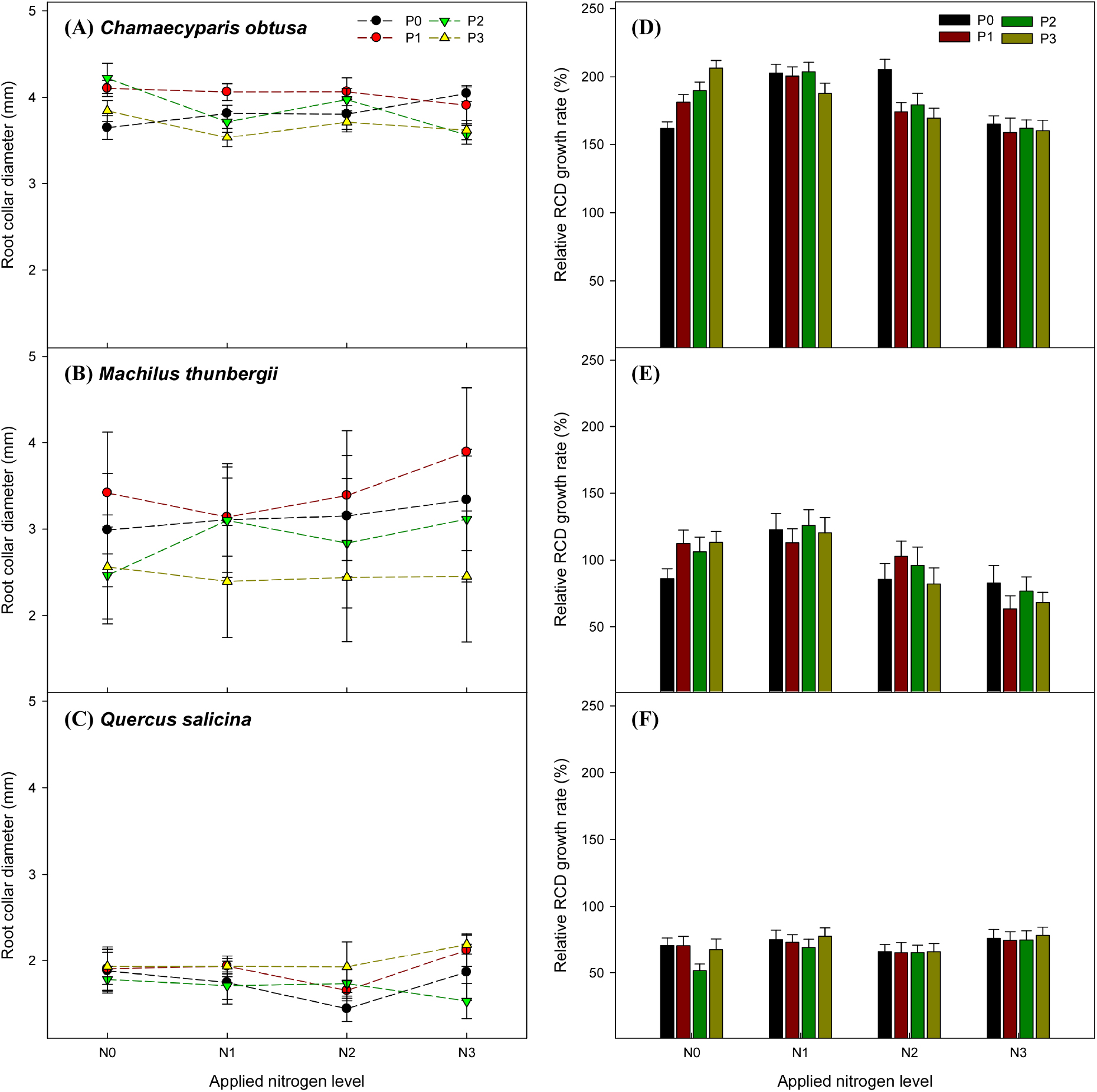
Fig. 1.
Root collar diameter (RCD) of (A) Chamaecyparis obtusa, (B) Machilus thunbergii, (C) Quercus salicina and relative RCD growth rate of (D) Chamaecyparis obtusa, (E) Machilus thunbergii, (F) Quercus salicina. The weight applied nitrogen levels were N0 (0 g·m-2), N1 (13.8 g·m-2), N2 (27.6 g·m-2), and N3 (41.4 g·m-2), while the weight applied phosphorus levels were P0 (0 g·m-2), P1 (6.1 g·m-2), P2 (12.2 g·m-2), and P3 (18.3 g·m-2).
질소 및 인 시비의 상호작용은 편백의 묘고 생장률에 유의한 영향을 미쳤으나, 후박나무와 참가시나무의 묘고 생장률에는 유의한 영향을 미치지 못하였다. 질소 시비는 세 수종 모두 묘고 생장에 통계적으로 유의한 영향을 미쳤지만(p < 0.001), 인 시비는 근원경 생장과 마찬가지로 묘고 생장에 유의한 영향을 미치지 못하였다. 질소 단일 처리에서는 편백과 후박나무는 질소 1배(N1P0)에서, 참가시나무는 질소 3배(N3P0)에서 가장 높았으며, 각각 대조구(N0P0)보다 약 1.4배, 1.4배, 1.1배 높았다(Fig. 2).
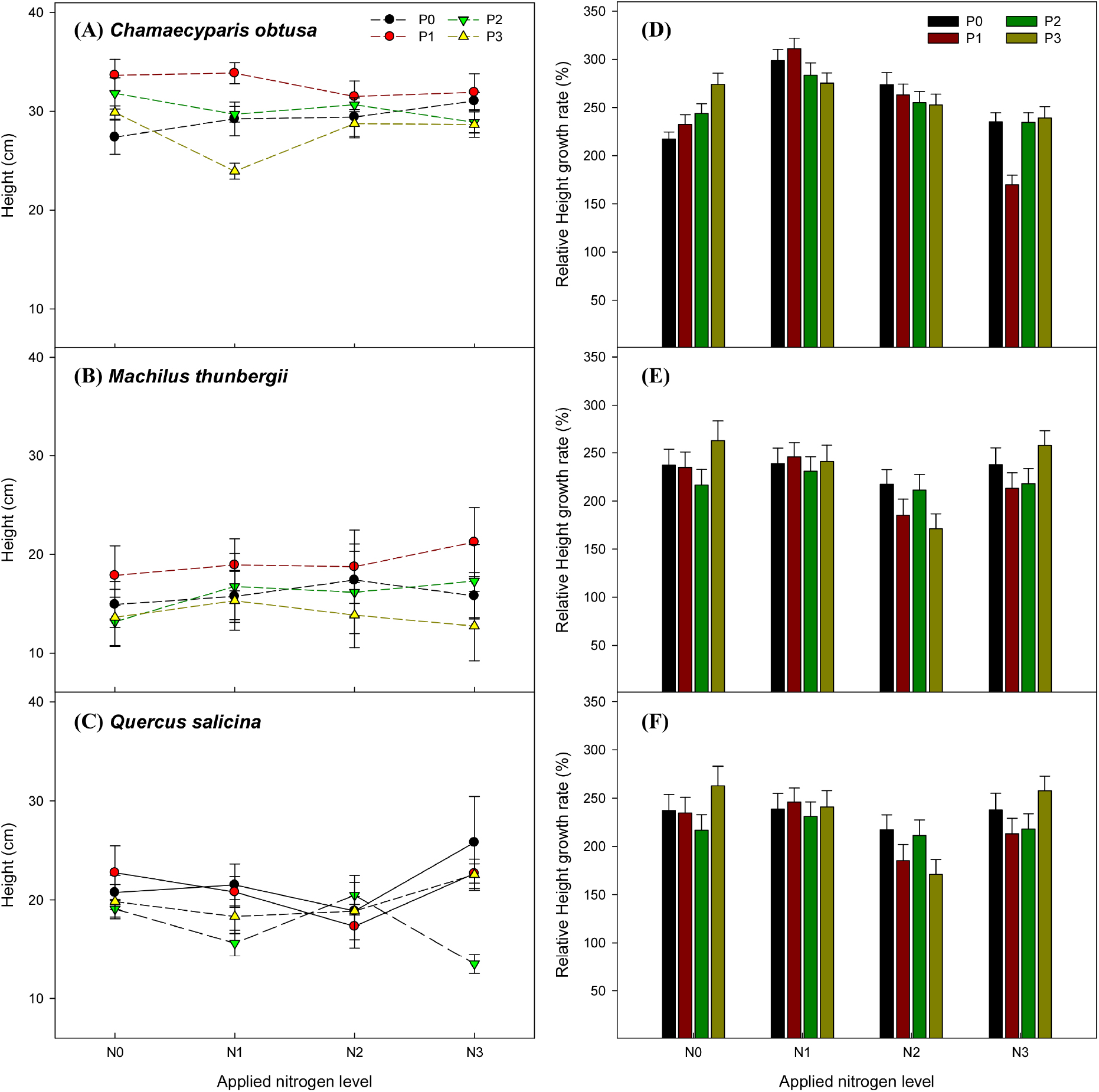
Fig. 2.
Height of (A) Chamaecyparis obtusa, (B) Machilus thunbergii, (C) Quercus salicina and relative height rate of (D) Chamaecyparis obtusa, (E) Machilus thunbergii, (F) Quercus salicina. The weight applied nitrogen levels were N0 (0 g·m-2), N1 (13.8 g·m-2), N2 (27.6 g·m-2), and N3 (41.4 g·m-2), while the weight applied phosphorus levels were P0 (0 g·m-2), P1 (6.1 g·m-2), P2 (12.2 g·m-2), and P3 (18.3 g·m-2).
Gojon 등(1991)은 질소 시비 농도가 증가함에 따라 잎과 줄기에서 질산 환원과 질산환원효소 활성이 증가하여 수목의 생장이 촉진된다고 보고하였다. 그러나 Sonneveld와 Voogt (2009)은 질소 시비가 적정 수준을 초과할 경우 오히려 수목의 생장이 감소할 수 있다고 언급하였다. 이를 바탕으로 볼 때, 각 수종의 근원경과 묘고 생장은 질소 시비가 증가할수록 무조건적으로 증가하는 것이 아니라, 특정 한계값이 존재한다고 판단된다.
본 연구 결과에서 인 시비의 유의한 영향은 없었지만, 질소 시비에 따라 근원경과 묘고 생장량이 증가함을 알 수 있었다. 하지만 각 수종별로 양분요구량과 흡수 특성이 다르기 때문에, 동일한 시비처리에도 수종에 따라 생장반응이 다르게 나타난 것으로 사료된다(Shin et al., 1999; Cho et al., 2011).
엽면적비(SLA)
질소시비는 편백을 제외한 나머지 두 수종의 SLA와 통계적으로 유의한 차이를 보이지 않았으며, 질소와 인의 복합처리, 인 시비 모두 세 수종의 SLA에 유의한 영향을 미치지 못하였다(Table 7). 본 연구에서. 편백의 SLA는 질소 시비에 따른 유의한 차이를 보였다(p < 0.05). 일반적으로 질소 시비는 엽면적과 SLA를 증가시킨다고 알려져 있지만(Knops and Reinhart, 2000; Chang, 2003), 본 연구에서는 질소 시비가 0배(N0P0), 1배(N1P0)일 때 SLA값이 크고, 2배(N2P0), 3배(N3P0)일 때 SLA 값이 감소하는 경향이 나타났다. 이는 식재 당년에 시비가 시행되어 광합성 산물이 잎보다는 뿌리 등에 분배되는 것과 관련된 것으로 사료된다(Canham et al., 1999). 이러한 현상은 질소 시비가 식물의 생장과 생리적 특성에 미치는 복합적인 영향을 반영하기 때문이다. 예를 들어, 질소 시비는 엽록소 함량과 광합성 능력을 증가시켜 생장을 촉진하지만, 과도한 질소 시비는 오히려 생장을 저해할 수 있다(Sonneveld and Voogt, 2009). 따라서 적절한 질소 시비 수준을 유지하는 것이 중요하다.
Table 7.
Specific leaf area (cm2·g-1) by species according to nitrogen and phosphorus fertilization.
| Species |
Nitrogen (g·m-2) | Phosphorus (g·m-2) | |||
| 0 | 6.1 | 12.2 | 18.3 | ||
| Chamaecyparis obtusa | 0 | 61.30 (0.19)z | 60.27 (0.26) | 59.91 (0.26) | 58.74 (0.36) |
| 13.8 | 61.96 (0.55) | 60.87 (0.35) | 57.28 (0.26) | 59.07 (0.54) | |
| 27.6 | 61.59 (0.18) | 60.97 (0.12) | 60.91 (0.20) | 59.80 (0.19) | |
| 41.4 | 61.31 (0.19) | 64.75 (1.97) | 59.78 (0.27) | 58.45 (0.20) | |
| Machilus thunbergii | 0 | 88.04 (0.95) | 86.01 (0.77) | 89.36 (1.72) | 100.97 (6.41) |
| 13.8 | 89.37 (0.83) | 85.18 (0.86) | 86.63 (0.79) | 87.64 (1.21) | |
| 27.6 | 91.84 (0.59) | 84.64 (0.32) | 84.47 (0.56) | 87.34 (1.66) | |
| 41.4 | 90.41 (0.80) | 85.33 (0.64) | 87.95 (1.82) | 88.96 (0.87) | |
| Quercus salicina | 0 | 98.43 (1.11) | 93.29 (1.12) | 97.03 (0.77) | 99.31 (0.65) |
| 13.8 | 97.37 (1.05) | 100.44 (1.07) | 99.11 (1.37) | 97.96 (0.94) | |
| 27.6 | 99.21 (1.15) | 97.20 (0.67) | 97.77 (1.25) | 100.44 (0.77) | |
| 41.4 | 94.29 (0.69) | 96.27 (0.70) | 96.51 (1.32) | 99.97 (0.49) | |
시비수준에 따른 광합성량
시비처리에 따른 광합성 반응의 결과를 세 수종의 광합성이 질소 시비와(p < 0.001), 인 시비에 따라(p < 0.05) 유의미한 차이가 있음을 보여준다. 또한 질소와 인 시비와 상호작용이 광합성에 유의미한 영향을 미침을 알 수 있다(Fig. 3). 적용된 시비 수준 아래에서 가장 광합성 값이 높은 4가지의 시비수준은, 편백에서 N1P0 > N2P1 > N0P0 > N1P2 순으로, 후박나무가 N2P3 > N3P3 > N0P3 > N1P0 순으로, 참가시나무는 N3P0 > N1P3 > N1P2 > N0P3 순으로 높게 나타났다. 인 비료를 처리하지 않은 경우, 편백과 후박나무는 질소 2배(N2P0)에서, 참가시나무는 4배(N4P0)에서 가장 높게 나타났으며, 질소 비료를 처리하지 않은 경우, 편백은 인 0배(N0P0)에서 후박나무와 참가시나무는 인 3배(N0P3)에서 가장 높게 나타났다.
Cho 등(2011)의 연구결과에서 토양에 부족한 질소와 인을 투입하면 엽록소 함량을 증가시키고 수목의 광합성이 촉진되지만 그 수준은 수종별로 다르다는 것을 알 수 있다. 따라서 질소와 인 시비 처리는 수목의 광합성 활성에 중요한 영향을 미치며, 최적의 시비 수준은 수종별 생리적 특성 따라 다르게 나타난다.
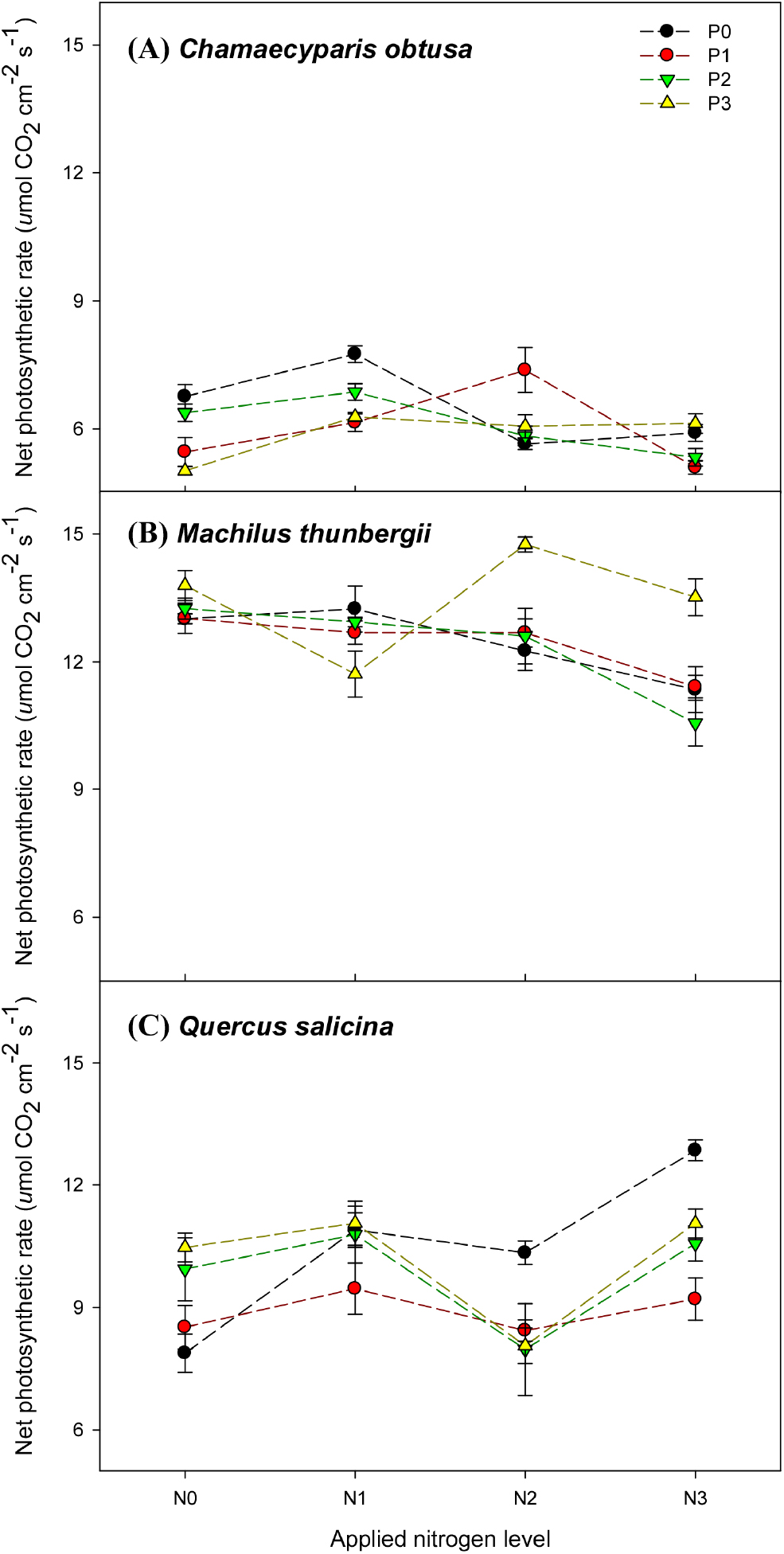
Fig. 3.
Photosynthesis according to fertilization of (A) Chamaecyparis obtusa, (B) Machilus thunbergii, (C) Quercus salicina. The weight applied nitrogen levels were N0 (0 g·m-2), N1 (13.8 g·m-2), N2 (27.6 g·m-2), and N3 (41.4 g·m-2), while the weight applied phosphorus levels were P0 (0 g·m-2), P1 (6.1 g·m-2), P2 (12.2 g·m-2), and P3 (18.3 g·m-2).
시비수준의 따른 잎 양분
세 수종 모두 잎의 질소 농도에 대해 인 시비와 질소 및 인 시비의 상호작용과는 통계적으로 유의하지 않았다.
편백과 후박나무의 경우, 질소 시비가 증가했을 때 잎의 질소 농도가 증가하는 경향을 보였으며, 통계적으로 유의함을 보였다(p < 0.05, Tables 8 and 9). 잎의 질소 농도가 가장 높게 나타난 처리구는 편백, 후박나무, 참가시나무 순으로 N3P2, N3P1, N3P1 이었고, 가장 낮게 나타난 처리구는 N0P2, N3P0, N2P2이다. 잎의 질소 농도가 가장 높게 나타난 처리구들의 근원경과 묘고 생장률을 살펴보면, 다른 처리구들에 비해 작다는 것을 알 수 있다. 이는 식물체의 생장이 활발하여 질소 농도가 바이오매스의 증가로 희석되었을 것으로 예상한다(Jarrell and Beverly, 1981).
Table 8.
Leaf nutrients properties after treatments about Chamaecyparis obtusa.
|
Properties (g·kg-1) |
Nitrogen (g·m-2) | Phosphorus (g·m-2) | |||
| 0 | 6.1 | 12.2 | 18.3 | ||
| N | 0 | 16.30 (0.46)z | 16.87 (0.50) | 16.29 (0.72) | 16.72 (0.26) |
| 13.8 | 18.90 (0.32) | 19.57 (0.75) | 19.02 (0.09) | 18.40 (0.31) | |
| 27.6 | 17.53 (0.17) | 17.83 (1.03) | 19.14 (0.62) | 19.01 (0.61) | |
| 41.4 | 17.88 (1.19) | 16.29 (0.72) | 19.21 (0.34) | 18.86 (0.37) | |
| P | 0 | 3.69 (0.08) | 3.65 (0.10) | 3.36 (0.07) | 3.31 (0.18) |
| 13.8 | 2.56 (0.39) | 2.82 (0.02) | 2.78 (0.03) | 2.55 (0.01) | |
| 27.6 | 2.67 (0.11) | 2.39 (0.22) | 2.32 (0.06) | 2.25 (0.00) | |
| 41.4 | 2.45 (0.09) | 2.57 (0.07) | 2.17 (0.18) | 2.07 (0.03) | |
| K | 0 | 14.57 (0.42) | 13.46 (0.67) | 14.18 (0.35) | 13.58 (0.85) |
| 13.8 | 13.29 (1.69) | 13.19 (0.94) | 12.41 (0.16) | 12.32 (0.51) | |
| 27.6 | 13.00 (0.62) | 12.89 (1.12) | 12.92 (0.83) | 13.02 (1.69) | |
| 41.4 | 12.78 (0.66) | 13.45 (0.35) | 12.48 (0.64) | 12.89 (0.86) | |
| Ca | 0 | 3.23 (0.16) | 3.28 (0.15) | 3.50 (0.21) | 3.03 (0.44) |
| 13.8 | 3.26 (0.15) | 3.04 (0.02) | 3.56 (0.01) | 3.34 (0.29) | |
| 27.6 | 3.66 (0.15) | 5.70 (1.04) | 3.77 (0.06) | 4.46 (0.68) | |
| 41.4 | 4.03 (0.47) | 3.97 (0.59) | 3.72 (0.41) | 4.51 (0.54) | |
| Mg | 0 | 1.96 (0.08) | 1.89 (0.02) | 1.96 (0.02) | 1.89 (0.08) |
| 13.8 | 1.96 (0.00) | 1.97 (0.00) | 2.14 (0.04) | 1.99 (0.08) | |
| 27.6 | 1.89 (0.10) | 1.94 (0.11) | 1.92 (0.11) | 1.99 (0.05) | |
| 41.4 | 1.85 (0.11) | 1.84 (0.02) | 2.09 (0.10) | 1.96 (0.01) | |
Table 9.
Leaf nutrients properties after treatments about Machilus thunbergii.
|
Properties (g·kg-1) |
Nitrogen (g·m-2) | Phosphorus (g·m-2) | |||
| 0 | 6.1 | 12.2 | 18.3 | ||
| N | 0 | 13.48 (0.68)z | 13.38 (0.31) | 14.67 (0.18) | 14.54 (0.40) |
| 13.8 | 12.80 (3.00) | 13.72 (0.23) | 14.96 (0.39) | 14.10 (0.60) | |
| 27.6 | 15.09 (0.50) | 14.92 (0.25) | 15.38 (0.46) | 15.18 (0.14) | |
| 41.4 | 16.21 (0.89) | 15.70 (0.46) | 14.21 (1.03) | 15.35 (0.48) | |
| P | 0 | 1.36 (0.17) | 1.44 (0.07) | 1.40 (0.07) | 1.35 (0.08) |
| 13.8 | 1.20 (0.03) | 1.29 (0.03) | 1.11 (0.03) | 1.08 (0.00) | |
| 27.6 | 0.96 (0.04) | 0.98 (0.10) | 0.96 (0.11) | 1.10 (0.23) | |
| 41.4 | 0.92 (0.00) | 1.06 (0.10) | 0.89 (0.11) | 1.03 (-) | |
| K | 0 | 11.88 (0.15) | 11.73 (0.60) | 11.53 (0.78) | 11.12 (0.48) |
| 13.8 | 10.28 (0.83) | 10.94 (0.83) | 10.29 (0.08) | 9.83 (0.63) | |
| 27.6 | 10.64 (0.05) | 9.92 (-) | 10.52 (0.77) | 10.34 (0.36) | |
| 41.4 | 11.31 (1.69) | 10.19 (0.51) | 10.67 (0.19) | 10.39 (0.17) | |
| Ca | 0 | 3.23 (0.16) | 3.28 (0.15) | 3.50 (0.21) | 3.03 (0.44) |
| 13.8 | 3.26 (0.15) | 3.04 (0.02) | 3.56 (0.01) | 3.34 (0.29) | |
| 27.6 | 3.66 (0.15) | 5.70 (1.04) | 3.77 (0.06) | 4.46 (0.68) | |
| 41.4 | 4.03 (0.47) | 3.97 (0.59) | 3.72 (0.41) | 4.51 (0.54) | |
| Mg | 0 | 2.18 (0.13) | 2.15 (0.01) | 2.35 (0.07) | 2.17 (0.11) |
| 13.8 | 1.99 (0.18) | 2.02 (0.07) | 2.01 (0.16) | 1.89 (0.10) | |
| 27.6 | 1.89 (0.11) | 2.06 (0.07) | 1.94 (0.07) | 1.77 (0.15) | |
| 41.4 | 1.84 (0.02) | 1.80 (0.02) | 1.73 (0.04) | 1.98 (0.17) | |
참가시나무 잎의 질소 농도는 시비처리의 영향이 통계적으로 유의하지 않았다(Table 10). 인 시비가 없을 때 질소 시비에 따른 질소 농도는 거의 차이가 없었지만, 나머지 처리에서 질소 시비가 증가할수록 질소 농도가 증가하였다.
Table 10.
Leaf nutrients properties after treatments about Quercus salicina.
|
Properties (g·kg-1) |
Nitrogen (g·m-2) | Phosphorus (g·m-2) | |||
| 0 | 6.1 | 12.2 | 18.3 | ||
| N | 0 | 17.60 (1.02)z | 17.66 (0.26) | 17.02 (0.11) | 16.52 (0.43) |
| 13.8 | 16.96 (0.19) | 16.26 (0.25) | 17.05 (0.23) | 17.84 (0.67) | |
| 27.6 | 17.04 (0.88) | 18.87 (0.06) | 17.33 (1.23) | 17.19 (0.23) | |
| 41.4 | 16.78 (2.37) | 20.00 (0.46) | 18.49 (0.28) | 17.06 (1.08) | |
| P | 0 | 1.23 (0.18) | 1.28 (0.06) | 1.20 (0.03) | 1.17 (0.10) |
| 13.8 | 1.18 (0.08) | 1.21 (0.01) | 1.16 (0.06) | 1.14 (0.06) | |
| 27.6 | 1.18 (0.07) | 1.11 (0.09) | 1.19 (0.03) | 1.28 (0.01) | |
| 41.4 | 1.34 (0.05) | 1.17 (0.10) | 1.20 (0.01) | 1.12 (0.01) | |
| K | 0 | 10.15 (0.59) | 11.13 (0.83) | 11.05 (0.50) | 11.32 (0.25) |
| 13.8 | 12.06 (0.31) | 12.30 (2.00) | 12.82 (2.17) | 10.12 (0.01) | |
| 27.6 | 11.01 (0.59) | 10.62 (0.50) | 10.68 (0.15) | 11.07 (1.52) | |
| 41.4 | 9.91 (1.98) | 9.87 (0.61) | 9.36 (0.18) | 9.28 (0.63) | |
| Ca | 0 | 7.36 (1.52) | 6.61 (0.01) | 6.50 (0.16) | 5.24 (0.04) |
| 13.8 | 5.95 (0.48) | 6.95 (0.99) | 5.59 (0.24) | 5.60 (0.13) | |
| 27.6 | 6.09 (0.14) | 4.38 (1.05) | 6.10 (0.71) | 5.23 (2.26) | |
| 41.4 | 7.54 (1.18) | 5.70 (0.14) | 5.32 (1.38) | 4.91 (1.15) | |
| Mg | 0 | 1.39 (0.09) | 1.47 (0.05) | 1.39 (0.18) | 1.28 (0.05) |
| 13.8 | 1.38 (0.05) | 1.37 (0.19) | 1.23 (0.06) | 1.22 (0.09) | |
| 27.6 | 1.28 (0.06) | 1.24 (0.14) | 1.34 (0.10) | 1.68 (0.05) | |
| 41.4 | 1.46 (0.11) | 1.32 (0.10) | 1.58 (0.17) | 1.58 (0.11) | |
편백과 후박나무 잎의 인 농도는 질소 시비가 증가할수록 감소하는 경향을 보였다(p < 0.001). 참가시나무의 경우, 질소 시비가 증가할수록 감소하다가 질소 3배량에서는 인 농도가 증가하는 모습을 보이며, 통계적으로 유의하다(p < 0.05). 이는 흡수과정 중 음이온의 길항작용 때문일 것이라고 판단된다(Mengel and Kirkby, 1987). 인 시비는 세 수종 모두 잎의 인 농도에 통계적으로 유의한 영향을 주지 않았다. 토양 내 인산 함량은 증가하였으나, 식물체의 인 흡수 속도가 느려 활용하지 못한 것으로 판단된다(Park et al., 1998)
편백과 후박나무 잎의 포타슘 농도는 N0P0에서, 참가시나무의 포타슘 농도는 N1P2에서 가장 높게 나타나며, 질소 시비가 증가할수록 감소하는 경향을 보이나 통계적으로 유의하지 않았다. 편백과 참가시나무 잎의 마그네슘 농도는 일정한 경향성을 보이지 않았으나, 후박나무 잎의 마그네슘 농도는 질소 시비가 증가할수록 감소하는 경향을 보였다(p < 0.001). 이는 질소와의 길항작용으로 인해 나타난 것으로 예상된다(Marschner, 2011).
Conclusion
본 연구는 질소와 인 시비 처리가 편백, 후박나무, 참가시나무의 생장, 광합성, 잎의 양분 특성에 미치는 영향을 조사하였으며, 묘목 생산 과정에서 수종에 따라 비료의 종류 및 시비량이 다르게 적용되어야 함을 보여주고 있다. 결과에서 질소 시비는 세 수종의 근원경과 묘고 생장, 광합성에 유의미한 영향을 미친 반면, 인 시비는 생장 특성에 유의미한 영향을 미치지 못하였다. 그러나 질소와 인 시비의 상호작용은 광합성 반응과 일부 양분 특성에 유의한 영향을 미치는 것으로 나타났다.
편백의 경우, 질소 시비가 2배량(N2P0)일 때 근원경과 묘고 생장이 가장 우수하였으며, 광합성 반응은 질소 시비가 1배량(N1P0)일 때 가장 높게 나타났다. 후박나무는 질소 1배량(N1P0)에서 가장 우수한 근원경 생장과 가장 높은 광합성을 보였다. 참가시나무는 질소 3배량(N3P0)에서 근원경 생장이 가장 우수하였으며, N3P0처리에서 가장 높은 광합성을 보였다. 이는 각 수종이 요구하는 최적 질소 시비 수준이 다르다는 것을 보여준다.
질소 시비는 편백과 후박나무의 잎 질소 농도를 증가시키는 경향을 보였으나, 과도한 시비는 생장을 저해하거나 양분의 희석 효과를 나타내었다. 후박나무 잎의 마그네슘 농도는 질소 시비가 증가할수록 감소하였으며, 이는 질소와 마그네슘 간 길항작용 때문으로 판단된다. 반면, 인 시비는 세 수종의 잎 인 농도에 유의미한 영향을 미치지 않았으며, 이는 토양 내 인산 함량은 증가했지만 식물체의 인 흡수 속도가 느려 활용되지 못했기 때문으로 보인다.
질소와 인 시비 처리는 광합성 반응에서도 중요한 역할을 하였다. 질소와 인 시비의 상호작용은 광합성량에 유의미한 영향을 미쳤으며, 이는 적절한 시비 수준을 설정하는 것이 중요함을 시사한다. 그러나 시비 수준이 과도할 경우, 생장 저하 및 양분 불균형이 발생할 수 있었다. 따라서, 최적의 시비 수준은 수종별 생리적 특성과 토양 조건을 종합적으로 고려하여 설정해야 한다.
본 연구의 결과는 질소와 인의 단일처리뿐만 아니라 복합처리의 효과를 검토함으로써, 묘목 생산 과정에서 시비수준(배율)의 조합이 생장에 미치는 영향을 구체적으로 설명할 수 있는 기초자료를 제공한다. 따라서 향후 연구에서는 복합처리의 세부 배율 설정과 장기적 효과를 평가하여 묘목 생산과 관련된 시비 전략을 정량화할 필요가 있다. 이를 통해 난대수종인 편백, 후박나무, 참가시나무의 적응성과 생산성을 높이고, 더욱 효율적인 우량 묘목 생산이 이루어질 수 있을 것으로 기대된다.
