

Introduction
Materials and Methods
Animals
Estrus synchronization and artificial insemination
Data set
Statistical analysis
Results
Effect of hormone treatment
Effect of inseminator
Effect of parity of the dam
Effect of AI year
Effect of AI season
Discussion
Conclusion
Introduction
Interest in the reproductive performance of cows continues to increase. In accordance with this trend, the 2023 public notice, “Livestock Improvement Goals” by the Ministry of Agriculture, Food and Rural Affairs (MAFRA), stated a goal to review the potential use of reproductive traits as future selection traits in Hanwoo cows by collecting and analyzing related data. Efforts of selecting superior cows by using genetic evaluation have continuously been made (MAFRA, 2023).
Reproductive traits are important economic traits in Hanwoo cows because they directly affect farm productivity and income. However, systematic investigation results are limited; this is mainly because the collection of large-scaled, accurate, and detailed reproductive information from Hanwoo cattle farms over a long period of time is challenging. Recently, the scale of Hanwoo farming and the number of full-cycle farms have increased. Hence, integrated production systems have expanded; planned breeding and marketing have become essential for improving productivity and management efficiency. Therefore, continuous efforts are required to improve the reproductive efficiency through field investigations and scientific analyses of the reproductive performance of cows.
Many reproductive indicators have been studied in dairy and beef cattle, including Hanwoo cattle. Services per conception (SPC) is a direct indicator of the number of artificial inseminations (AIs) required for a cow to become pregnant. A low SPC indicates a high insemination efficiency. As the number of services increases, the costs of AI, semen, and labor also rise; the open period may lengthen, leading to economic losses. Therefore, SPC can be used as an objective reproductive trait to evaluate the reproductive efficiency of cows (Chang et al., 2006; Kim and Jeong, 2018).
Generally, reproductive traits (including SPC) possess low heritability and are strongly affected by environmental factors. Hence, it is important to identify measurable environmental factors in the target population and establish an appropriate statistical model to accurately evaluate the genetic ability of each animal (Choi et al., 2005; 2018; Eum et al., 2016).
We investigated SPC in Hanwoo cows and analyzed the various environmental factors affecting it. This is done by comparing SPC associated with AI following hormone-based estrus synchronization and SPC associated with AI following the detection of natural estrus to investigate potential differences in environmental effects. We aimed to provide basic evidence that can be used for future analyses and genetic evaluations of reproductive traits in Hanwoo cows.
Materials and Methods
Animals
This study utilized AI records from the Hanwoo Research Center located in Daegwallyeong-myeon, Pyeongchang-gun, Gangwon-do Province, Korea. The records were collected from 2020 to the first half of 2024; only cows that became pregnant and calved normally with a single calf were included. Cases that required five or more inseminations to achieve pregnancy and calving were excluded because they may possess reproductive disorders.
After quality control, 1,141 records were considered usable for analysis. The records included 672 from the line-breeding population and 469 from the base population. The line-breeding Hanwoo population (LBH) has been maintained at the Hanwoo Research Center since 2009 using selected proprietary sires, whereas the general Hanwoo population (GH) was established using proven Korean bulls (Jin et al., 2023; 2024).
The Hanwoo Research Center performs AI twice per year (in the first and second halves). The two populations were inseminated and calved in the same cowhouses during each insemination season and were managed under the same feeding and management system. All procedures were approved by the Institutional Animal Care and Use Committee of the National Institute of Animal Science (Approval No. 2020-449).
Estrus synchronization and artificial insemination
The estrus synchronization protocol, which is applied to all specimens, is described below (Fig. 1). Each cow received an intravaginal progesterone device containing 1.9 g of progesterone (controlled internal drug release, CIDR; Zoetis, New Zealand) at 09:00 irrespective of their estrus cycle stage. Simultaneously, 2 mL of gonadotropin-releasing hormone (GnRH; 100 µg gonadorelin acetate/mL; Fertagyl, Germany) was intramuscularly injected in the neck.
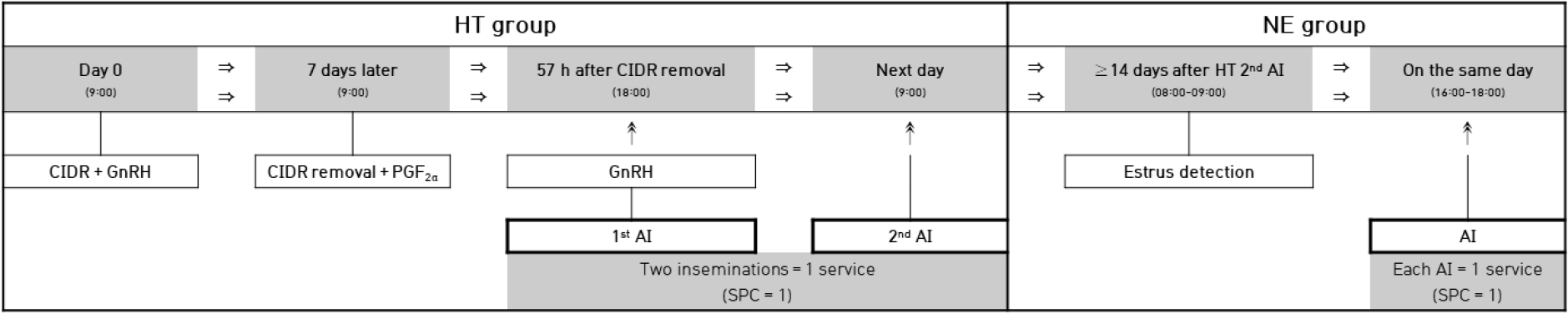
Fig. 1.
Estrus synchronization and artificial insemination protocol used in the HT and NE groups. HT, hormone-treated estrus synchronization; NE, natural estrus; CIDR, controlled internal drug release; GnRH, gonadotropin-releasing hormone; PGF2α, prostaglandin F2 alpha; AI, artificial insemination; SPC, services per conception.
Seven days later, the CIDR was removed at 09:00; 5 mL of prostaglandin F2α (PGF2α; 5 mg dinoprost tromethamine/mL; Lutalyse, Zoetis, Belgium) was injected. Ovulation was induced by an intramuscular injection of 2 mL of GnRH at 57 h post-CIDR removal. The first and second AI were conducted at the time of GnRH administration and at 09:00 the next morning, respectively.
Frozen semen was provided in 0.5 mL straws containing 18 - 20 million sperm each. Before AI, straws were thawed in 37℃ water for 40 s, loaded into an AI gun with a protective sheath, and used for insemination. Trained technicians performed all inseminations; the details followed Kang et al. (2024) and Jin et al. (2024).
Data set
Cows that calved after the synchronization protocol described above were assigned to the hormone-treated group (HT). In the HT group, SPC was calculated per synchronization cycle (Fig. 1); therefore, two inseminations performed within the same synchronization cycle (18:00 and 09:00) were counted as one service. The AI date was recorded as the date of the second insemination (09:00), and that synchronization cycle was counted as SPC = 1.
Cows that returned to estrus after the second insemination received additional AI. Estrus behavior was observed from 08:00 to 09:00; one AI was performed from 16:00 to 18:00 on the same day. These inseminations were recorded separately for the natural estrus group (NE). In the NE group, SPC was calculated per insemination (Fig. 1); therefore, each insemination following a detected estrus event was counted as a separate service. To minimize the residual effects of previous hormone treatment and to distinguish the two management systems more clearly, only additional AI records occurring at least 14 days after the second insemination were included in the NE analysis.
Inseminators were classified within each group to evaluate the differences in SPC among them. Only inseminators with ≥ 20 AIs were included in HT, whereas those with ≥ 10 AIs were included in NE. Consequently, six inseminators were analyzed in each group.
Parity groups were defined as primiparous (parity 1) cows and multiparous cows with parity ≥ 2. For additional parity analysis, HT was divided into eight categories (parities 1 - 7 and ≥ 8); NE was divided into five categories (parities 1 - 4 and ≥ 5). The AI season was classified into the first (March - August) and second (September - February of the following year) halves of the year.
Statistical analysis
All analyses were conducted using the R software (R Core Team, 2024). The main objective of this study was to test the differences in SPC across various environmental factors. The SPC was analyzed using a linear model to compare mean differences across factors; fixed effects were evaluated using analysis of variance (ANOVA). In this framework, the estimated mean SPC represents the average number of services required per conception within each level of the evaluated factor, and the corresponding p-values indicate whether mean SPC differed significantly among factor levels. When the ANOVA indicated significant differences, Duncan’s new multiple range test based on the mean square error was applied for multiple comparisons. Because SPC is an integer-valued outcome with a restricted range, sensitivity analyses were performed using a generalized linear model (with a Poisson family and log link) and an ordinal logistic regression model. Because the primary aim of this study was to compare average SPC across environmental factor levels in a straightforward and interpretable manner, the linear model was selected as the main analytical approach. As the conclusions regarding the significance of the main effects were consistent across all models, the results from the linear model are primarily presented for intuitive interpretation.
Results
Effect of hormone treatment
There was no significant difference in SPC between LBH and GH. The mean SPC ± standard deviation (SD) was 1.55 ± 0.848 for LBH and 1.47 ± 0.752 for GH (p = 0.097) (Table 1). In contrast, the SPC was significantly lower (p < 0.001) in HT than in NE (Table 2). The mean SPC ± SD was 1.38 ± 0.756 in HT and 2.06 ± 0.785 in NE. Based on these results, further analyses of the environmental factors affecting SPC were performed using HT and NE rather than the other two populations (LBH and GH). Within each group, the SPC did not differ significantly between LBH and GH (data not shown).
Table 1.
Comparison of the number of SPC between LBH and GH groups.
| Population | N | Mean ± SD | SE | Min | Max | p-value |
| LBH | 672 | 1.55 ± 0.848 | 0.031 | 1 | 4 | 0.097 |
| GH | 469 | 1.47 ± 0.752 | 0.037 | 1 | 4 |
Table 2.
Comparison of SPC between HT and NE groups.
| Group | N | Mean ± SD | SE | Min | Max | p-value |
| HT | 906 | 1.38b ± 0.756 | 0.025 | 1 | 4 | < 0.001 |
| NE | 235 | 2.06a ± 0.785 | 0.050 | 1 | 4 |
Effect of inseminator
In HT, the mean SPC of the inseminators who performed at least 20 AIs ranged from 1.19 to 1.52. Differences among inseminators were statistically significant (p = 0.026); however, the mean differences were small and the confidence intervals overlapped, suggesting limited practical importance. In NE, the mean SPC of the inseminators who performed at least 10 AIs ranged from 1.71 to 2.26; no significant differences were detected among the inseminators (p > 0.05) (Table 3).
Table 3.
Comparison of the number of SPC among AI technicians between HT and NE groups.
Effect of parity of the dam
When the dams were classified as primiparous or multiparous, HT exhibited no significant differences. In HT, the mean SPC ± standard error (SE) was 1.41 ± 0.05 for primiparous dams and 1.37 ± 0.029 for multiparous dams (p > 0.05).
In NE, the mean SPC ± SE was 1.82 ± 0.082 for primiparous dams, which was significantly lower than 2.29 ± 0.056 for multiparous dams (p < 0.001) (Table 4).
Table 4.
Comparison of the number of SPC between primiparous and multiparous cows in HT and NE groups.
Parity was further classified within each group (Table 5). In HT, when parity was divided into eight categories (1 - 7 and ≥ 8), mean SPC values ranged from 1.32 to 1.50; no significant differences were observed among parity classes (p > 0.05). In NE, when parity was divided into five categories (1 - 4 and ≥ 5), the mean SPC ranged from 1.82 to 2.44. Dams with parities 1 and 2 exhibited lower SPC than dams with parity ≥ 3 (p < 0.001). The lowest mean SPC ± SE was observed in parity 1 (1.82 ± 0.082); SPC tended to increase with parity, showing a clear difference between parities 1 and 2 and parity ≥ 3.
Table 5.
Comparison of the number of SPC according to parity between HT and NE groups.
Effect of AI year
Across the AI years from 2020 to 2024, SPC did not differ significantly in either group (p > 0.05). In HT, the annual mean SPC ranged from 1.24 to 1.62; in NE, it ranged from 1.85 to 2.22 (Table 6).
Table 6.
Comparison of the number of SPC by year of AI.
Effect of AI season
When the SPC was compared between the first (March - August) and second (September - February) halves of the AI seasons, no significant seasonal effect was observed in HT (p > 0.05). In NE, mean SPC ± SE was 2.17 ± 0.065 in the first half of the season, which was significantly higher than 1.96 ± 0.078 in the second half of the season (p < 0.05) (Table 7).
Table 7.
Comparison of the number of SPC by season of AI.
Discussion
We investigated environmental factors affecting SPC in Hanwoo cows. We analyzed SPC by separating cows inseminated after estrus synchronization using hormone treatment from those inseminated after naturally detected estrus.
Previous studies on SPC in Hanwoo cows reported a wide range of values. Choi et al. (2005) investigated reproductive traits at five Hanwoo farms and reported a mean SPC ± SD of 1.78 ± 1.04, with no significant correlation between body condition score and SPC. Kim et al. (2009) reported that the SPC ± SE in primiparous Hanwoo cows from the Chungcheongbuk-do Province was 1.6 ± 0.2, which was slightly higher than that in cows with parity ≥ 2; however, the difference was not significant. Kim et al. (2014) reported SPC values of 1.4 - 1.7 and emphasized the importance of accurate estrus detection and reproductive record management. Eum et al. (2016) reported a mean SPC ± SD of 1.21 ± 1.21, whereas Kwon et al. (2019) reported a mean SPC ± SD of 1.411 ± 0.803; both studies found significant differences by parity and season.
Sasaki et al. (2016) reported a mean SPC ± SD of 1.60 ± 0.96 and found significant effects of herd size, season, parity, and calving-to-first service interval (CFSI) on SPC in Japanese Black cattle. They also reported interactions between herd size and season, herd size and parity, and herd size and CFSI. In small and medium herds, the SPC was higher for inseminations performed in winter and spring, whereas no seasonal differences were observed in large herds. Irikura et al. (2018) reported a mean SPC ± SE of 1.6 ± 0.02 in nulliparous heifers and of 1.7 ± 0.01 in parous cows; cows with SPC of 1 - 3 had a significantly higher conception rate than cows with SPC ≥ 4. Setiaji and Oikawa (2020) reported a mean SPC ± SD of 1.76 ± 1.08 in Japanese Black heifers and estimated a very low heritability for SPC (0.019). They also reported a negative correlation between SPC and non-return rate (-0.363) and a strong positive correlation between SPC and first-service conception rate (0.874).
In the present study, the mean SPC ± SD was 1.55 ± 0.848 in LBH and 1.47 ± 0.752 in GH. These values were higher than the mean reported by Eum et al. (2016) (1.21) and lower than that reported by Choi et al. (2005) (1.78); they were within the range observed in previous studies of Hanwoo cows. However, when the cows were divided into HT and NE, the mean SPC in HT was 1.38, which was significantly lower than 2.06 in NE (p < 0.001). Direct comparisons of these group-specific SPC values with previous studies are limited, because most earlier reports presented overall SPC values without distinguishing between hormone-treated estrus synchronization and natural estrus-based insemination.
The relatively higher SPC observed in the NE group may be associated with the difficulty of precise AI timing under natural estrus detection compared with the scheduled insemination used in HT. However, this difference should be interpreted with caution because SPC was calculated differently between the two groups. In the HT group, one service represented one synchronization cycle including two timed inseminations, whereas in the NE group, each insemination after a detected estrus event was counted as a separate service. Therefore, the lower SPC observed in HT may reflect not only improved reproductive management associated with hormonal synchronization, but also, in part, the different counting rules used for SPC calculation. In addition, differences in AI protocols, recording systems, and environmental conditions among herds and studies should also be considered when interpreting these results.
Our results showed that in HT, inseminator, parity, AI year, and AI season did not significantly affect SPC. In contrast, in NE, SPC was significantly lower in primiparous cows than in multiparous cows, whereas cows with parity ≥ 3 exhibited significantly elevated SPC (p < 0.001). Although the mean SPC did not differ significantly by year (p > 0.05), it was significantly higher in the first half of the season than in the second half (p < 0.05). This pattern partly agrees with previous reports, indicating that parity and season contribute to variations in SPC.
To improve the reproductive traits in Hanwoo cows, including SPC, correlations among reproductive traits and genetic correlations with economically important traits, such as carcass and growth traits, must be evaluated. This approach will allow for a careful assessment of how improvements in reproductive performance are linked to productivity and overall economic outcomes.
Hormone-based estrus synchronization and timed AI (TAI) was developed to address challenges in estrus detection and low reproductive efficiency; they are now widely used in both dairy and beef cattle (Bó and Baruselli, 2014; Colazo and Mapletoft, 2014). Modern programs, such as Ovsynch and its presynchronization variants (e.g., Presynch-Ovsynch and Double-Ovsynch), combine GnRH and PGF2α to enable AI at a fixed time without estrus detection and improve reproductive outcomes, including conception at the first TAI after calving (Pursley et al., 1995; Souza et al., 2008). At the farm level, these programs have been reported to shorten the interval to reinsemination and improve conception and pregnancy rates, with particular benefits in cows with weak estrus expression or in high-producing dairy cows (Bó and Baruselli, 2014).
A key advantage of estrus synchronization is that it reduces the labor and time required for estrus detection while allowing all animals to be inseminated at the exact planned time. In contrast, AI based on natural estrus depends strongly on accurate estrus detection and correct insemination timing, which are major determinants of reproductive success (Colazo and Mapletoft, 2014; Diskin, 2018). Therefore, the elevated SPC observed in the NE of the present study likely reflects missed optimal insemination timing due to incomplete or inconsistent estrus observations. This suggests that errors in estrus detection can directly reduce insemination efficiency; it also indicates that estrus synchronization or fixed TAI (FTAI) can help ensure appropriate insemination timing and improve reproductive performance, particularly in large herds.
In the NE of the present study, primiparous cows exhibited a significantly lower SPC than multiparous cows. Generally, multiparous cows often exhibit lower fertility than primiparous cows owing to postpartum uterine recovery, changes in the uterine environment, and increased metabolic stress, such as a negative energy balance in high-producing cows (Rizos et al., 2010). Experimental evidence also suggests that the reproductive tract environment in postpartum cows (oviduct and uterus) may be less supportive of early embryo development than that in primiparous cows, which could explain the elevated SPC in multiparous cows (Rizos et al., 2010). However, when hormone-based synchronization and TAI are applied, combinations of progesterone, GnRH, and PGF2α can control key stages of the estrus cycle (follicular growth, luteolysis, and ovulation), reduce dependence on estrus detection, and reduce variation among animals, thereby increasing uniform reproductive outcomes (Colazo and Mapletoft, 2014). TAI can also minimize or remove estrus detection requirements and has been widely used as a herd-level strategy for improving reproductive efficiency (Baruselli et al., 2017).
SPC can vary greatly owing to farm- and animal-level factors, such as herd size, season, parity, and CFSI. Management systems within herds (e.g., resynchronization programs) and inseminator effects can also contribute to differences in conception-related performance (Sasaki et al., 2016; Setiaji and Oikawa, 2020). Additionally, reproductive traits generally exhibit low heritability (approximately 0.02 - 0.04) (Chang et al., 2006; Berry et al., 2014). Hence, for traits strongly influenced by environmental factors, such as SPC, genetic effect estimates can be biased unless the model properly accounts for contemporary groups (e.g., herd - year - season) and major management factors. This highlights the need for the systematic modeling of environmental effects (Van Bebber et al., 1997; Choi et al., 2005; Eum et al., 2016).
Our findings also suggest that hormone treatment for estrus synchronization may be a major management factor, explaining the variation in SPC. FTAI enables AI at a scheduled time without estrus detection (Bó and Baruselli, 2014); the success of TAI programs can differ depending on protocol design (e.g., Presynch-Ovsynch) and synchronization response, leading to differences in pregnancy rate or pregnancy per AI (Colazo and Mapletoft, 2014). Hence, future genetic analyses and breeding studies on SPC may need to consider the hormone treatment status, either as a fixed effect or by defining contemporary groups. This approach helps distinguish environmental effects and allows for an improved estimation of the genetic reproductive ability of each animal (Van Bebber et al., 1997; Colazo and Mapletoft, 2014). Overall, the results of the present study provide basic information for future analyses of reproductive traits and the development of genetic evaluation models for Hanwoo cows.
Conclusion
This study quantified SPC in Hanwoo cows and examined the major environmental sources of variation using AI records. SPC did not differ between LBH and GH; however, it differed substantially in the insemination setting. Specifically, the SPC was lower for inseminations performed under hormone-based estrus synchronization than for those performed following the detected natural estrus. Within the synchronization setting, inseminator differences were small yet statistically detectable; parity, AI year, and season were not significant. In contrast, under natural-estrus-based AI, SPC increased with parity and was higher in the first half of the season (March - August) than in the second half (September - February), indicating that parity- and season-related variations were more evident when insemination depended on estrus detection than when insemination depended on hormone-based synchronization.
Overall, the insemination setting (hormone treatment status) emerged as a major management factor associated with SPC variation in this dataset. These findings provide baseline information for future reproductive studies in Hanwoo cattle and suggest that hormone treatment status should be considered when modeling SPC, either as a key management factor or through contemporary group definitions, to better account for environmental variation in subsequent genetic and breeding analyses.
