

Introduction
Materials and Methods
고온기 딸기 재배시 트레할로스 엽면 살포 실험
열 충격 전사인자 발현 분석 실험
Results
실험 기간 동안의 온도 변화
트레할로스 처리는 ‘설향’ 딸기의 생육 특성에 영향을 미치지 않음
트레할로스 처리는 사계성 딸기 ‘w-33’의 생육 특성에 영향을 미치지 않음
트레할로스 처리는 열 충격 전사인자 FvHsfA2a의 발현을 지속시킴
Discussion
Conclusion
Introduction
딸기(Fragaria × ananassa Duch.)는 장미과에 속하는 초본성 과채류로, 생식용뿐만 아니라 디저트 장식, 음료 및 잼과 같은 가공용으로도 널리 이용된다. 2024년 기준 국내딸기 생산액은 약 1조 5,584억 원으로, 채소작물 중 가장 높은 점유율을 차지하고 있다(MAFRA, 2026). 국내에서는 딸기가 주로 시설하우스에서 촉성 및 반촉성 재배 방식으로 재배되며, 주로 겨울과 봄철에 과실 생산이 이루어진다. 딸기는 저온성 작물로, 영양 번식에 의해 증식되며, 육묘는 주로 4월에서 9월까지 시설 내에서 이루어진다. 그러나 여름철 고온으로 인해 양질의 묘 생산이 어려운 문제가 있다(Nam et al., 2015; Lee et al., 2016). 또한, 사계성 딸기를 여름철 고온기에 재배할 경우 고온 장해로 인해 생육 및 과실 품질이 저하되는 문제가 발생한다(Lee et al., 2017). 특히, 여름철 고온다습한 시설 환경은 탄저병, 시들음병, 역병 등의 병해 발생을 촉진한다(Lee et al., 2018). 따라서 고온기 딸기 재배 시 스트레스 및 병해를 경감할 수 있는 기술 개발이 필요하다.
트레할로스(α-D-glucopyranosyl-[1–1]-α-D-glucopyranoside)는 두 분자의 포도당으로 이루어진 비환원성 이당류로, 식물에서 신호 분자로 작용하는 것이 알려져 있다(Paul et al., 2008; Smeekens et al., 2010). 또한, 고온, 저온, 건조, 염 스트레스와 같은 비생물적 스트레스(Krasensky and Jonak, 2012)뿐만 아니라 다양한 생물적 스트레스 반응과 생리적 과정의 조절에도 관여하는 것으로 보고되었다(Singh et al., 2011). 외인성 트레할로스 처리는 식물의 생물적 및 비생물적 스트레스 저항성을 향상시키는 데 큰 잠재력을 가지며, 이 과정에는 여러 생리적 메커니즘이 관련되어 있다(Ali et al., 2012; Luo et al., 2022).
식물은 고온 스트레스에 반응하여 신호전달 과정을 통해 물리적 및 생화학적 변화를 유도하며, 분자 수준에서는 고온 스트레스로부터 보호하는 유전자의 발현이 변화한다(Shinozaki and Yamaguchi-Shinozaki, 2007; Hasanuzzaman et al., 2013). 예를 들어, 열 충격 전사인자(heat shock transcription factors, HSFs)는 고온 스트레스를 포함한 다양한 환경 스트레스 조건에서 식물 보호 관련 유전자 발현을 조절하여 식물의 스트레스 저항성을 향상시키는 역할을 한다(Schramm et al., 2008; Yoshida et al., 2010; Scharf et al., 2012; Hwang et al., 2014; Samadi et al., 2019). 따라서 딸기에서 트레할로스 처리가 비생물적 스트레스 반응에 미치는 영향을 평가하기 위해 열 충격 전사인자의 유전자 발현을 분석할 필요가 있다.
이처럼 신호 생성, 전달 및 방어 반응을 통해 식물의 생물적 및 비생물적 스트레스 저항성을 유도하는 물질은 환경 스트레스 경감 및 병해충 관리에 활용될 수 있는 친환경적인 제재로 주목받고 있다(García-Mier et al., 2013). 트레할로스는 최근 식품, 화장품 및 의약품 원료로도 많이 활용되고 있으며(Ohtake and Wang, 2011), 가지과 작물을 대상으로 친환경 재배에 활용하기 위한 연구가 진행된 바 있다(Mori et al., 2023). 특히, 껍질째 섭취하는 딸기의 경우 친환경 제재의 사용이 요구되지만, 딸기 재배에서 트레할로스 처리 효과에 대한 연구는 거의 없다. 국내에서도 트레할로스의 식물 대사 및 재배상 이용에 대한 연구가 적은 편이다. 저자들의 선행 연구에서는 여름철 고온기에 사계성 딸기에 트레할로스를 장기간 토양 관주 처리한 결과, 딸기의 지상부 생육 및 과실 생육 특성, 수량에는 유의한 변화가 없었으나, 팁번 증상과 시들음병 발생이 유의하게 감소하는 경향을 보였다(Lee et al., 2025). 이러한 결과는 트레할로스가 배지의 수분 보유력 및 전기전도도(electrical conductivity, EC)를 조절하여 고온 스트레스를 경감하는 데 기여할 가능성을 시사한다(Mohanan et al., 2023).
따라서 본 연구에서는 여름철 고온기 딸기 육묘 및 사계성 딸기 재배 시 트레할로스를 장기간 엽면 살포하였을 때 생육에 미치는 영향을 평가하고자 하였다. 또한, 고온 스트레스 하에서 딸기의 열 충격 전사인자 발현과 트레할로스 처리의 영향을 분석하였다. 본 연구를 통해 딸기 재배 시 고온 스트레스 경감에 대한 트레할로스의 관여 여부를 확인하고, 스트레스 경감제로서의 활용 가능성을 검토하고자 한다.
Materials and Methods
고온기 딸기 재배시 트레할로스 엽면 살포 실험
본 연구는 2023년 여름철(6월 16일 - 9월 7일) 세종특별자치시에 위치한 우리종묘(36°33.913′ N, 127°22.351′ E) 플라스틱 온실에서 수행되었다. 온실 내 온도는 온도/습도 기록계(174H, Testo, Germany)를 이용하여 1시간 간격으로 측정하였다.
트레할로스 엽면 살포 실험은 고설 배드에서 양액(Tank Farming A/B, Agroad, Korea)으로 재배 중인 딸기 식물체를 대상으로 수행되었다. 실험은 두 개의 조건에서 진행되었다. 첫 번째 실험에서는 육묘 중인 ‘설향’ 딸기를 대상으로 하였으며, 두 번째 실험에서는 우리종묘에서 선발한 사계성 딸기 ‘w-331’을 여름 재배 조건에서 평가하였다. 두 실험 모두 난괴법 3반복으로 배치하였으며, 처리구당 10개체를 배치하였다.
트레할로스 엽면 살포 처리는 대조구(물) 및 10, 30 mM의 트레할로스 수용액을 사용하였으며, 압축 분무기를 이용하여 잎의 앞·뒷면에 고르게 도포하였다. 처리 일정은 6월 16일부터 7월 14일까지는 1주일에 1회, 이후 9월 7일까지는 2주일에 1회씩 총 9회 수행하였다.
생육 조사는 농촌진흥청 딸기 생육 조사 방법을 참고하여 수행하였다. 측정 항목은 엽장, 엽폭, 초장, 관부 직경, 엽수로 설정하였으며, 엽장 및 엽폭은 50 cm 알루미늄 자를 이용하여 측정하였다. 초장은 지제부에서 가장 긴 잎까지의 길이를 측정하였으며, 관부 직경은 디지털 버니어 캘리퍼스(SD600-150W, Sincon, China)를 이용하여 가장 넓은 부분과 가장 좁은 부분을 측정한 후 평균값을 사용하였다.
‘설향’ 모주 및 자묘의 생육 조사는 처리 후 83일 경과 시점(9월 7일)에 수행하였다. 모주의 생육은 처리당 30주를 대상으로 조사하였으며, 자묘의 생육은 처리당 45주를 수확하여 평가하였다. 자묘의 엽면적은 엽면적 측정기(LI-3100, LI-COR, USA)를 사용하여 분석하였다. 생체중과 건조중은 지상부 및 지하부로 구분하여 측정하였으며, 건조중 측정을 위해 70℃에서 3일간 건조하였다. 팁번 증상 및 병징 발생 정도는 0 - 5까지 6단계(0: 없음, 1: 약간, 3: 중간, 5: 심함)로 구분하여 평가하였다.
사계성 딸기 ‘w-331’의 생육 및 과실 특성 조사는 처리 후 34일(7월 20일) 시점에서 수행하였다. SPAD 값은 측정기(SPAD-502, Minolta, Japan)를 이용하여 정단으로부터 세 번째 잎에서 세 지점을 측정한 후 평균값을 사용하였다. 과실은 수확 후 과고, 과폭, 당도를 측정하였다. 과고 및 과폭은 디지털 버니어 캘리퍼스를 이용하여 가장 긴 부분과 가장 짧은 부분을 측정한 후 평균값을 사용하였다. 당도는 비파괴 당도계(P-100, Sunforest, Korea)를 이용하여 측정하였다. 팁번 및 병징 발생 정도는 ‘설향’과 동일한 기준으로 조사하였다.
통계분석은 R (R Core Team, 2022)을 사용하였다. 농도별 트레할로스 처리가 딸기에 미치는 효과가 있는지 확인하기 위해 일원 분산분석을 진행하였고 ‘agricolae’ 패키지를 설치하여 사후검정으로 던컨의 다중 검정(Duncan’s multiple range test)을 진행하였다.
열 충격 전사인자 발현 분석 실험
본 실험에서는 사계성 딸기 ‘w-331’의 세 번째 잎을 시료로 사용하였다. 트레할로스 처리가 잎 전체에 균일하게 전달될 수 있도록 물꽂이 방법을 적용하였다. 증류수 또는 30 mM 트레할로스 용액이 담긴 유리병에 잎을 꽂아 72시간 동안 처리한 후, 42℃로 설정된 식물 생장상에 배치하여 스트레스를 유도하였다. 스트레스 처리 후 0, 0.5, 1시간 경과 시점에서 샘플을 채취하고, 액체질소로 즉시 동결하였다.
액체질소로 동결한 딸기 잎을 막자와 막자사발을 이용하여 마쇄한 후, Apure™ Plant RNA (Apbio, Korea) 키트를 이용하여 total RNA를 분리하였다. 합성된 cDNA는 ReverTra Ace -α-™ (Toyobo, Japan) 키트를 사용하여 생성하였다.
PCR 증폭은 SimpliAMP Thermal Cycler (Thermo Fisher Scientific, USA)를 사용하여 수행하였으며, 18S rRNA 유전자를 내재 대조군으로 사용하였다. PCR 반응 혼합물은 preMix AccuPower® (Bioneer, Korea)를 이용하였다. PCR 반응 조건은 95℃에서 15초, 64℃에서 20초, 72℃에서 40초를 35회 반복한 후, 72℃에서 7분간 최종 신장 반응을 수행하였다. PCR 분석에 사용된 프라이머 염기서열은 Hu 등(2015)을 참고하여 설정하였으며, 프라이머 정보는 Table 1에 제시하였다.
Results
실험 기간 동안의 온도 변화
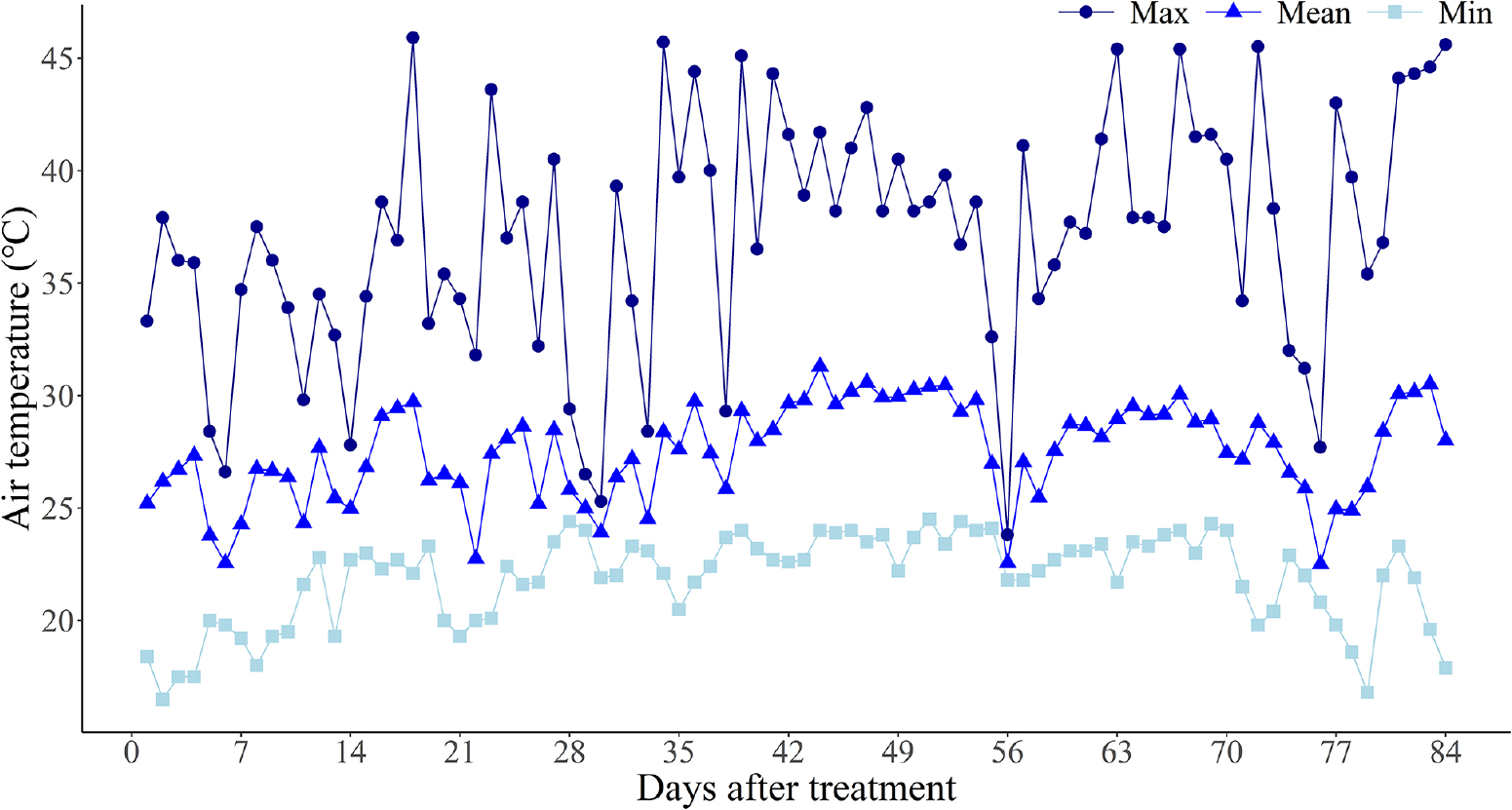
Fig. 1은 실험 기간(2023년 6월 17일 - 9월 7일) 동안 세종특별자치시에 위치한 우리종묘 온실 내부의 기온 변화를 나타낸다. 월별 평균 기온은 6월 25.7℃, 7월 27.6℃, 8월 28.1℃, 9월 28.3℃로 측정되었으며, 최고 기온은 각각 37.9℃, 45.9℃, 45.0℃, 45.6℃ 였다. 실험 기간 중 최고 기온이 40℃ 이상인 일수는 26일이었고, 35℃ 이상인 일수는 57일이었다. 일반적으로 딸기는 17 - 20℃ 의 온도에서 최적 생육을 보이는 저온성 작물로, 35℃ 이상의 온도에서는 생육이 저해되고, 40℃ 이상에서 3일 이상 노출될 경우 런너 생육이 심각하게 저해되는 것으로 알려져 있다(Kadir et al., 2006). 본 실험이 수행된 온실은 최고 기온이 45℃를 상회하는 등 일반적인 재배 한계 온도를 훨씬 초과하는 환경이었으며, 이에 따라 딸기 식물체는 생리적 방어 기작을 넘어서는 심각한 수준의 고온 스트레스를 받았을 것으로 판단된다.
트레할로스 처리는 ‘설향’ 딸기의 생육 특성에 영향을 미치지 않음
Table 2와 Table 3은 트레할로스 처리 후 83일 경과(9월 7일) 시점에서 ‘설향’ 딸기의 모주 및 자묘 생육 특성을 비교한 결과이다. 지속적인 트레할로스 엽면 살포(10 mM 및 30 mM)를 시행한 후 대조구와 처리구 간의 초장, 관부직경, 엽수, 엽장 및 엽폭의 차이를 비교하였으나, 유의미한 차이는 관찰되지 않았다. 또한, 자묘 수 역시 대조구와 처리구 간 차이가 없었다(Fig. 2). 자묘의 엽면적(Fig. 3), 지상부 및 지하부의 생체중과 건조중 역시 처리구 간 차이를 보이지 않았다(Fig. 4). 팁번 및 병징 발생 정도를 비교한 결과, 모주에서 자묘보다 다소 높은 증상이 나타났지만, 처리 간 차이는 없었다(Tables 2 and 3).
Table 2.
Growth effect of foliar spraying with 0, 10, and 30 mM trehalose on mother plant of ‘Sulhyang’ strawberry.
| Treatmentx |
Plant height (cm) |
Crown diameter (mm) |
No. of leaves |
Leaf length (mm) |
Leaf width (mm) |
Tip burn occurrence degreey |
Disease symptom occurrence degreey |
| Control | 32.1 ± 3.7az | 12.8 ± 1.8a | 10.3 ± 1.3a | 88.2 ± 13.1a | 70.3 ± 6.9a | 2 | 2 |
| Tre-10 | 33.7 ± 3.4a | 13.0 ± 1.5a | 10.0 ± 1.2a | 94.4 ± 14.4a | 71.8 ± 7.0a | 2 | 2 |
| Tre-30 | 32.9 ± 4.5a | 12.8 ± 1.2a | 10.7 ± 1.7a | 93.0 ± 15.0a | 75.0 ± 8.3a | 2 | 2 |
Table 3.
Effects on strawberry (cv. Seolhyang) daughter growth by trehalose foliar spray treatments in nursery bed during high temperature period.
| Treatmentx |
Plant height (cm) |
Crown diameter (mm) |
No. of leaves |
Leaf length (mm) |
Leaf width (mm) |
Tip burn occurrence degreey |
Disease symptom occurrence degreey |
| Control | 27.8 ± 1.16az | 7.4 ± 0.6a | 3.1 ± 0.3a | 82.3 ± 8.48a | 60.6 ± 3.4a | 1 | 1 |
| Tre-10 | 27.6 ± 2.39a | 7.3 ± 0.7a | 3.1 ± 0.4a | 80.1 ± 11.7a | 59.4 ± 5.7a | 1 | 1 |
| Tre-30 | 27.8 ± 1.77a | 7.2 ± 0.7a | 3.0 ± 0.3a | 79.7 ± 8.83a | 60.3 ± 5.5a | 1 | 1 |
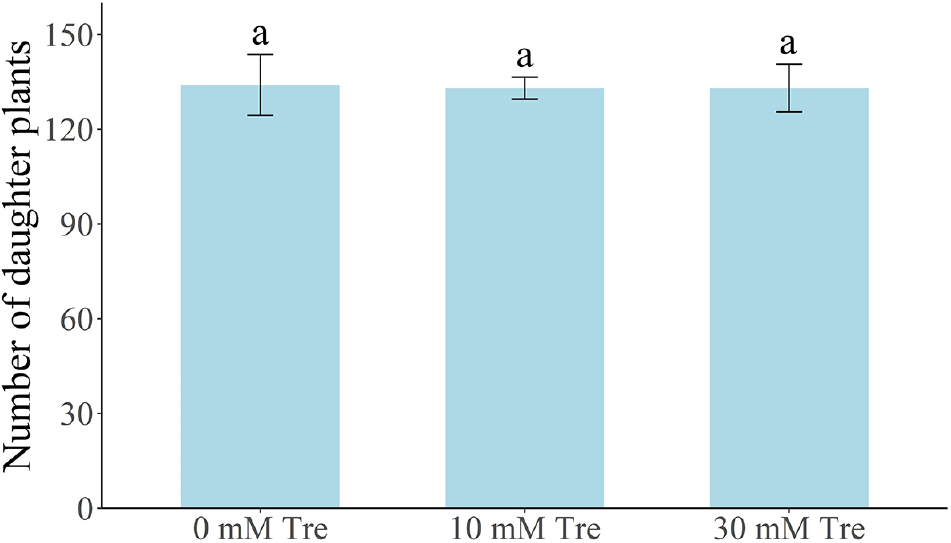
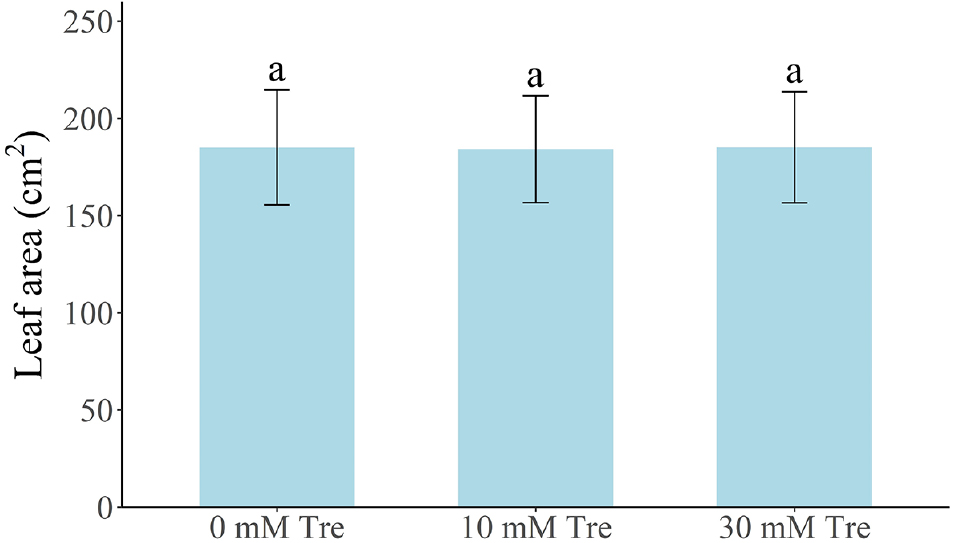
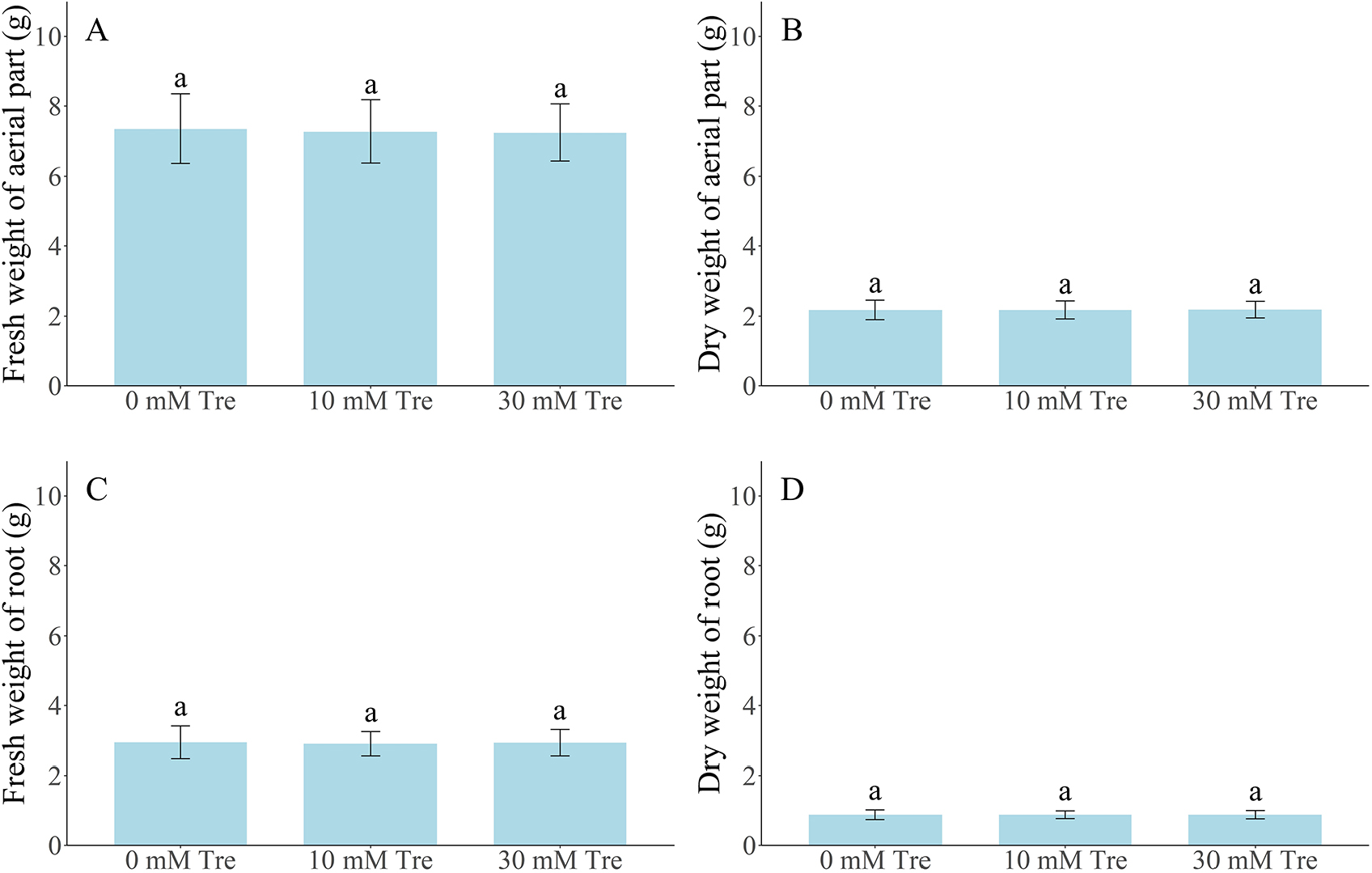
Fig. 4.
Fresh (A and C) and Dry weight (B and D) of ‘Sulhyang’ strawberry daughter plant following foliar spraying with 0, 10, and 30 mM trehalose at 83 days after treatment. Error bars indicate standard deviations (n = 45). Small letters represent significant differences; p ≤ 0.05 (Duncan’s multiple test).
트레할로스 처리는 사계성 딸기 ‘w-33’의 생육 특성에 영향을 미치지 않음
Table 4는 트레할로스 처리 34일 후(7월 20일) 사계성 딸기 ‘w-331’의 생육 특성을 조사한 결과이다. 대조구와 트레할로스 엽면 살포 처리구 간 초장, 관부직경, 엽수, 엽장 및 엽폭에서 유의적인 차이는 나타나지 않았다. 또한, 1차 수확한 과실의 과고, 과경, 당도 및 무게에서도 처리구와 대조구 간 유의미한 차이가 관찰되지 않았다(Table 5). 팁번 및 병징 발생 정도는 ‘설향’보다 ‘w-331’에서 다소 높게 나타났으나, 트레할로스 처리에 따른 유의적인 차이는 없었다(Table 4).
Table 4.
Effects on four-season strawberry (‘w-331’) growth by trehalose foliar spraying treatments during high temperature period.
| Treatmentx |
Height (cm) |
Crown diameter (mm) |
No. of leaves |
Leaf length (mm) |
Leaf width (mm) |
SPAD value |
Tip burn occurrence degreey |
Disease symptom occurrence degreey |
| Control | 20.5 ± 1.4az | 20.0 ± 1.3a | 8.9 ± 1.1a | 113.2 ± 4.9a | 90.1 ± 7.0a | 43.0 ± 4.7a | 2 | 3 |
| Tre-10 | 20.1 ± 1.6a | 19.7 ± 2.0a | 8.9 ± 1.1a | 113.7 ± 5.4a | 90.0 ± 8.0a | 42.3 ± 3.6a | 2 | 3 |
| Tre-30 | 20.0 ± 1.2a | 19.9 ± 1.8a | 9.0 ± 1.1a | 114.0 ± 3.7a | 88.7 ± 8.2a | 43.0 ± 4.5a | 2 | 3 |
Table 5.
Effects on fruit characteristics of four-season strawberry (‘w-331’) by trehalose foliar spraying treatments during high temperature period.
| Treatmenty |
Fruit height (mm) |
Fruit diameter (mm) |
Fresh weight (g) |
Soluble solids (°Brix) |
| Control | 51.3 ± 3.7az | 48.9 ± 3.8a | 18.1 ± 1.3a | 7.9 ± 0.4a |
| Tre-10 | 50.9 ± 3.4a | 49.3 ± 3.5a | 17.9 ± 1.2a | 8.0 ± 0.3a |
| Tre-30 | 51.1 ± 3.3a | 49.4 ± 3.1a | 18.0 ± 1.2a | 8.0 ± 0.3a |
트레할로스 처리는 열 충격 전사인자 FvHsfA2a의 발현을 지속시킴
본 연구에서는 트레할로스 처리가 딸기 잎에서 열 충격 전사인자 FvHsfA2a의 발현에 미치는 영향을 semi-qRT-PCR을 이용하여 분석하였다. Hu 등(2015)에 따르면, FvHsfA2a는 2배체 딸기(Fragaria vesca)에서 열 스트레스에 의해 가장 강하게 유도되는 열 충격 전사인자로, 42℃ 고온 처리 30분 만에 발현이 증가한 후 빠르게 감소하는 특성을 보인다. 본 연구에서도 42℃ 처리 30분 후 FvHsfA2a의 발현이 강하게 유도되는 것을 확인하였다.
주목할 점은, 30 mM 트레할로스 용액을 3일간 처리한 잎에서 FvHsfA2a의 유도 발현 양이 대조구(증류수 처리)보다 높았으며, 발현이 더 오랫동안 지속되었다(Fig. 5). 이는 트레할로스가 딸기 세포 내에서 고온 스트레스 신호 전달을 강화할 수 있음을 보여준다. 단, 이 결과는 통제된 생장상 내의 수경 재배 조건에서 얻어진 것으로, 복합적인 환경 요인이 작용하는 실제 온실 토양 재배 환경과는 반응의 차이가 있을 수 있다. 따라서 이 유전자 발현 결과는 트레할로스가 딸기의 스트레스 반응 기작에 관여할 수 있다는 잠재적 가능성을 확인한 것에 의의를 둔다.

Discussion
본 연구에서는 여름철 고온기에 딸기 재배 시 트레할로스의 스트레스 경감 효과를 평가하고, 효과적인 활용 방안을 탐색하였다. 연구 결과, 트레할로스의 엽면 살포는 ‘설향’ 딸기의 육묘 및 사계성 딸기 ‘w-331’의 생육과 과실 특성에 유의한 영향을 미치지 않았으며, 팁번 및 병해 발생률에서도 차이가 나타나지 않았다. 하지만 장기간 트레할로스 엽면 살포에 따른 약해 증상 또한 관찰되지 않아, 고농도(30 mM) 처리의 안전성은 확인되었다.
본 실험에서 트레할로스 처리에 따른 생육 증진 효과가 미미했던 원인은 다음 두 가지 측면에서 고찰해 볼 수 있다. 첫째, 극한의 고온 환경이다. 본 실험 기간 중 온실 내 최고 기온은 45.9℃에 달했으며, 40℃ 이상인 날이 26일이나 지속되었다. 이는 저온성 작물인 딸기가 생존할 수 있는 임계 온도를 크게 초과한 조건이다. 트레할로스가 삼투 조절제나 항산화제로 작용하여 세포를 보호한다 하더라도(Paul et al., 2008; Ali et al., 2012), 식물체 전체의 생리적 시스템이 붕괴될 수준의 고온 피해를 단독 엽면 처리만으로 극복하기에는 한계가 있었을 것으로 판단된다. 둘째, 엽면 살포 방식에 따른 흡수 효율의 문제이다. 트레할로스와 같은 당류는 잎의 기공과 큐티클층을 통해 흡수될 수 있으나(Fernández and Eichert, 2009), 고온 건조한 환경에서는 트레할로스 이 잎 표면에서 빠르게 건조되거나 식물의 기공이 닫혀 흡수율이 저하될 수 있다. 본 연구의 FvHsfA2a 발현 분석 결과는 트레할로스가 세포 내로 흡수될 경우 스트레스 신호 전달을 강화할 수 있음을 보여주었으나, 실제 온실 실험에서는 엽면 살포된 트레할로스가 유효 농도 이상으로 식물체 내에 충분히 흡수되지 못해 내성 증진 효과로 이어지지 못했을 가능성이 있다.
트레할로스의 효과는 처리 방법에 따라 달라질 수 있다. Mori 등(2023)은 가지과 작물에서 트레할로스 엽면 살포는 생육에 영향이 없었으나, 토양 관주 처리 시에는 주두 신장을 촉진하여 고온 피해를 경감시켰다고 보고하였다. 본 연구는 엽면 살포 방식으로만 진행되었기에 토양 관주 효과를 직접 검증하지는 못하였으나, 선행 연구 결과를 고려할 때 향후에는 엽면 살포 시 전착제(surfactant)를 첨가하여 흡수율을 높이거나, 뿌리를 통한 흡수를 유도하는 관주 처리(irrigation) 방식을 적용하여 고온 스트레스 경감 효과를 재평가할 필요가 있다. 또한, 본 연구에서는 일반적인 생육 지표(초장, 엽수 등)만을 조사하였으나, 향후 연구에서는 수분 이용 효율(water use efficiency, WUE), 엽록소 형광 반응, 자묘의 런너 활력 등 스트레스 반응에 더 민감한 생리적 지표를 활용한 정밀 분석이 요구된다.
종합하면, 트레할로스는 딸기 재배 시 약해 없이 사용할 수 있는 안전한 소재이며, 분자 수준에서 고온 스트레스 반응을 조절할 잠재력을 가지고 있다. 그러나 여름철 온실과 같은 극한 고온 조건에서 실질적인 재배적 이점을 얻기 위해서는 단순 엽면 살포를 넘어선 처리 방법의 최적화(농도, 흡수 증진제, 관주 처리 등)가 선행되어야 할 것이다.
Conclusion
본 연구는 여름철 고온기 딸기 재배 시 트레할로스의 친환경적 스트레스 경감 소재로서의 활용 가능성을 탐색하였다. ‘설향’ 및 ‘w-331’ 품종에 대한 장기간 트레할로스 엽면 살포 결과, 식물체에 약해(phytotoxicity)는 유발하지 않았으나, 극한의 고온 환경(최고 45.9℃) 하에서 생육 및 과실 품질의 유의한 증진 효과는 관찰되지 않았다. 반면, 통제된 조건에서의 유전자 분석 결과 트레할로스 처리가 열 충격 전사인자 FvHsfA2a의 발현을 증진시키는 것으로 나타나, 분자 수준에서의 스트레스 신호 조절 가능성은 확인되었다. 이러한 결과는 트레할로스의 엽면 살포 방식이 극한 고온 조건에서 식물체 내 유효 흡수 농도 도달에 한계가 있음을 시사한다. 따라서 향후 트레할로스를 효과적인 고온 경감제로 활용하기 위해서는 식물체 흡수 효율을 높일 수 있는 처리 기술(전착제 혼용, 토양 관주 등) 개발과 더불어, 다양한 고온 강도에서의 효과 검증 연구가 필요하다.
