

Introduction
Materials and Methods
연구 방법
연구대상지
LANDIS-II 입력 자료 구축
식생 자료 수집
Results and Discussion
LANDIS-II 구동 결과
식생 자료 수집 결과
연구대상지 식생 변화 비교 분석
Conclusion
Introduction
산불은 산림생태계의 구조와 기능을 근본적으로 변화시키는 주요 교란 요인으로, 생물 다양성 감소와 토양 특성 변화, 수문 순환의 교란 등 복합적인 생태계 영향을 초래한다(Donato et al., 2012; McLauchlan et al., 2020). 우리나라의 최근 10년(2015 - 2024) 평균 산불 발생 건수는 546건, 피해 면적은 약 4,003 ha에 달하며, 기후변화의 영향으로 발생 빈도와 강도가 증가하는 추세이다(KFS, 2025). 이러한 산불의 강도와 발생 양상은 생태계 변화를 좌우하는 주요 요인으로 작용하며, 식생 구조의 변화와 향후 산불의 확산 양상과 밀접하게 연관된다(Cui and Perera, 2008). 결과적으로 산림의 자생적 회복 능력을 저하시켜 기존 우점종의 쇠퇴와 새로운 수종의 확산을 촉진하며, 장기적인 식생천이 과정을 불명확하게 만든다(Miller et al., 2009).
산불 피해지 복원은 자연복원과 인공복원으로 구분된다(NIFoS, 2022). 자연복원은 생태계의 자생적 회복 능력과 천이 과정을 반영하여 인간의 개입을 최소화하는 접근으로, 교란 이후 환경조건과 종 간 상호작용을 통해 식생이 자율적으로 회복되도록 유도한다. 인공복원은 단기간 내 인위적 개입을 통해 경관을 회복하는 방식으로, 식재나 종자 파종 등을 통해 식생구조를 조성하는 방식이다(NIFoS, 2022). 복원 방식의 차이는 산불 이후 식생의 회복 과정에도 영향을 미치며, 종조성의 변화와 구조적 다양성의 발달은 장기적인 천이 과정을 결정하는 핵심 요인으로 작용한다(Chen et al., 2014). 이와 같이 산불과 식생 변화는 상호작용 관계를 이루며, 식생 회복 양상은 향후 교란에 대한 생태계의 저항성과 회복력을 평가하는 중요한 지표로 해석될 수 있다(Sturtevant et al., 2009; Poorter et al., 2021). 이러한 관점에서 산불 이후 식생천이를 정량적으로 분석하는 연구는 단순한 경관 복원 수준을 넘어 교란 체계 내에서 생태계 안정성과 회복 메커니즘을 규명하는 복원 생태학적 의의를 가진다(Johnstone et al., 2016). 장기적 식생 변화의 정량적 연구는 산불 피해지에 생태적으로 적합하고 지속가능한 식생복원계획에 과학적 근거를 제공할 수 있다(Xi et al., 2008).
우리나라는 1996년 고성 산불과 2000년 동해안 산불 이후 장기 모니터링을 통해 피해지의 식생천이 과정을 분석해 왔으나(NIFoS, 2016), 대부분의 연구는 특정 시점의 식생 자료를 기반으로 한 기술적 분석에 머물러 있다(Hwang et al., 2012; Byeon and Yun, 2018; Choung et al., 2024; Kim et al., 2024a; 2024b). 현장 조사, 원격탐사 접근만으로는 산불 이후의 장기적·공간적 식생천이 과정을 정량적으로 규명하기 어렵다는 한계가 있다.
국외에서는 모니터링 기반 접근법의 한계를 보완하기 위해 산림경관모델을 병행하여 장기적 식생 변화를 규명하고 있다(Barbosa et al., 2014; Laflower et al., 2016; Skre, 2020; Simon, 2021). 모델을 활용한 접근은 관측 자료 보완 수준을 넘어, 산불 이후 생태계가 어떤 과정을 거쳐 회복하고 안정 상태로 이행하는지를 탐색할 수 있는 복원생태학적 분석 틀로 확장되고 있다(He et al., 2002; Xi et al., 2008). 복원생태학은 교란 이후 생태계의 회복 과정과 안정성, 그리고 복원 잠재력의 평가를 주요 관점으로 하며, 산림경관모델은 이를 정량적으로 평가할 수 있는 유용한 수단으로 기능한다(Perera et al., 2015). 특정 환경 조건에서 효과적인 복원계획을 판단하는 근거를 제시하며(Crouzeilles et al., 2016; 2017), 산불 이후 복원 개입의 시기와 강도를 결정하는 데 활용될 수 있다(Beschta et al., 2004; Moreira et al., 2011). 장기 예측을 통해 복원 시나리오를 비교·평가하여 산불 이후 관리 계획 수립의 의사결정 지원 도구로 활용 가능하다(Xi et al., 2008; Chazdon and Guariguata, 2016). 특히, LANDIS-II (Landscape Disturbance and Succession) 모델은 기후, 토양, 지형, 종 특성과 같은 환경 요인을 반영하여 수종별 생장·경쟁·천이 과정을 장기적으로 모의할 수 있는 산림경관모델로, 산불, 병해충, 벌목 등의 교란 요인에 대한 산림의 구조적 변화를 예측하는 데 활용되고 있다(Scheller et al., 2007; De Bruijn et al., 2014). 국내에서도 LANDIS-II의 적용 사례가 점차 보고되고 있으나(Choi et al., 2019; Cho et al., 2020; Moon et al., 2022), 대부분 기후변화 시나리오 혹은 특정 교란 유형에 한정되어 있으며, 산불 이후 식생천이 과정을 정량적으로 예측한 연구는 부족한 실정이다.
따라서 본 연구는 2000년 동해안 산불로 피해를 입은 자연복원지를 대상으로 LANDIS-II 모델을 적용하여 25년간(2000 - 2025)의 식생천이 과정을 예측하고, 예측 결과를 현장 조사 자료와 비교·분석함으로써 모델의 국내 적용 가능성과 한계를 검토하였다. 이를 통해 산불 피해지의 자연복원 이후 식생천이 양상을 정량적으로 분석하고, 향후 생태적 식생복원계획 수립을 위한 기초자료를 제공하고자 한다.
Materials and Methods
연구 방법
식생천이 예측을 위해 LANDIS-II 모델의 PnET-Succession (Photosynthesis and EvapoTranspiration Succession) 모듈을 활용하였다. 본 연구에서는 2000년 동해안 산불로 인해 모든 식생이 초기 천이 단계(1년생)로 초기화된 것으로 설정하고 2000년부터 2025년까지 25년간의 식생천이를 모의하였다. 모델의 출력값은 각 시간 단계에서 생체량이 최대인 수종 하나만을 우점 수종으로 표현하므로(Suárez-Muñoz et al., 2021), 구동 결과에 나타난 우점종 분포는 생체량 기준 우점 수종을 의미한다. 예측된 식생 구조와 수종 분포는 현장 조사 자료와 비교하여 모델이 국내 산불 피해지의 천이 과정을 얼마나 재현할 수 있는지 분석하였다.
연구대상지
연구대상지는 1996년 4월 고성 산불, 2000년 4월 동해안 산불이 발생한 강원도 고성군 죽왕면 인정리 산1 일대로 북위 38°18′36″ - 38°19′22″, 동경 128°28′44″ - 128°29′44″에 위치한다(Fig. 1). 대상지의 전체 면적은 82 ha이며, 이 중 70 ha는 우리나라 최초로 산불 이후 산림청에 의해 자연복원 연구지역으로 지정된 곳이다(KFS, 1997; Choung et al., 2000). 이러한 최초의 자연복원 시도의 효과성을 측정하기 위해 국립산림과학원은 고정조사구 2곳을 설치하여 식생 회복, 야생동물, 토양 등에 대한 장기 모니터링을 현재까지 지속적으로 수행하고 있다(NIFoS, 2016). 인위적 간섭을 최소화한 상태로 관리되고 있어, 산불 이후 식생천이 과정을 규명하기에 적합한 연구대상지로 판단된다. 해발고도는 최고 175 m, 최소 45 m이며, 태백산맥과 동해안의 영향을 받는 중부동안형 기후에 속해 일반적인 온대성 기후를 보인다(Goseong County, 2025). 2000년 기준 연평균 기온은 12℃이며, 연평균 강수량은 1,345.2 mm이다.
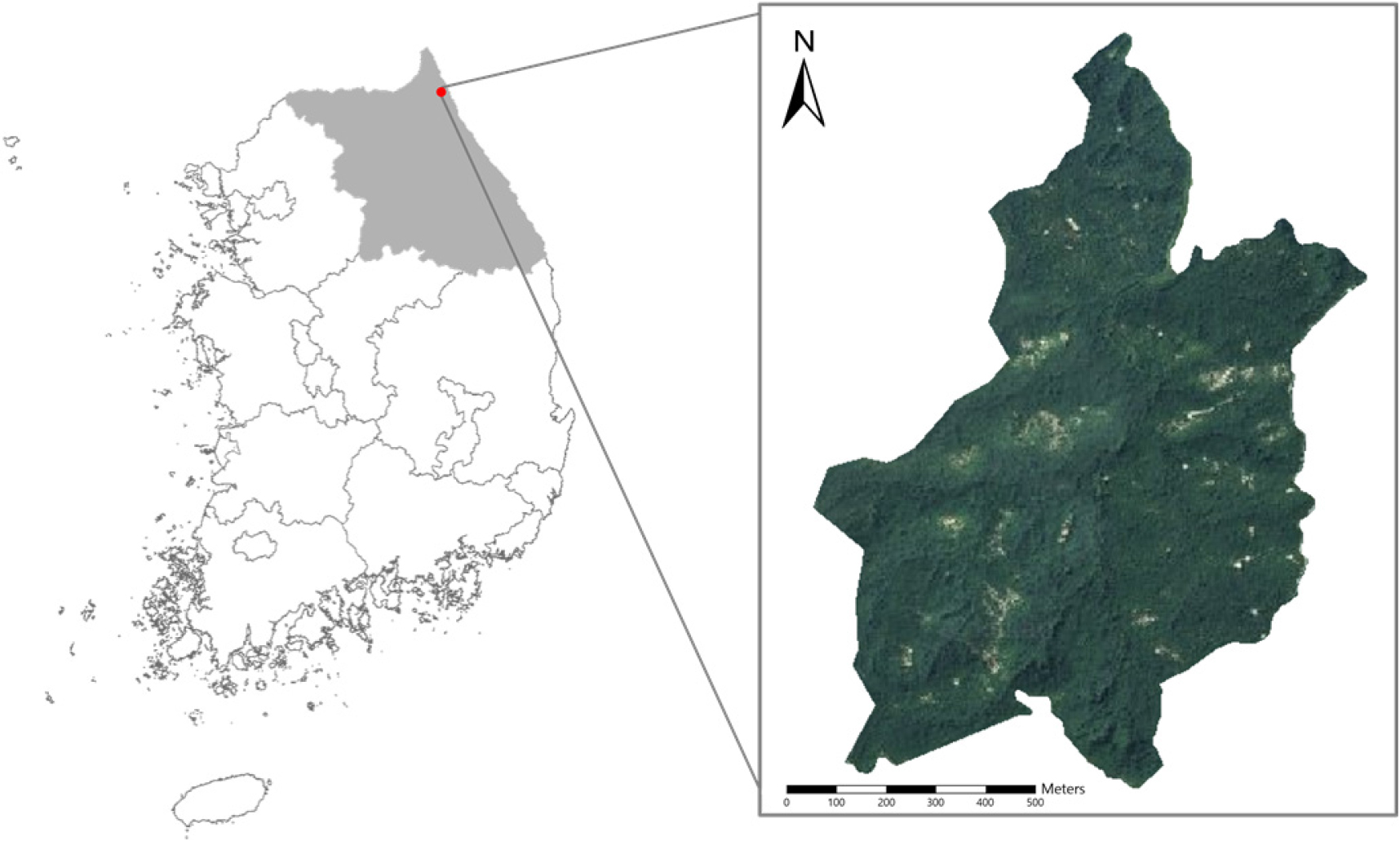
LANDIS-II 입력 자료 구축
LANDIS-II 구동에 필요한 핵심 입력 자료는 총 4가지로 구성되며, 생태역 지도(ecoregion map), 초기 식생지도(initial communities map), 기후자료(climate Data), 매개변수(parameters)가 있다. 연구대상지를 100 m2 정사각형 격자로 나누어 총 105개의 격자가 생성되었고, 각 격자마다 상기 자료를 입력하여 분석에 필요한 데이터를 구축하였다(Table 1).
Table 1.
Input data for LANDIS-II.
| Input data | Data type | Source | Resolution |
| Ecoregion map | Annual mean temperature | KMA (2000) | - |
| DEM | NGII (2024) | 1 : 5,000 | |
| Forest soil map | KFS (2003) | 1 : 25,000 | |
| Initial communities map | Vegetation map | ME (1990) | 1 : 50,000 |
| Digital forest type map | KFS (1993) | 1 : 25,000 | |
| Ecological survey of the Goseong wildfire-affected area | KFS (1997) | - | |
| Goseong wildfire white paper | Goseong County (1997) | - | |
| Climate data | Synoptic meteorological observations | KMA (2025) | - |
| PAR | Choi et al. (2021) | - | |
| CO2 | Gustafson et al. (2023) | - |
생태역 지도(ecoregion map)
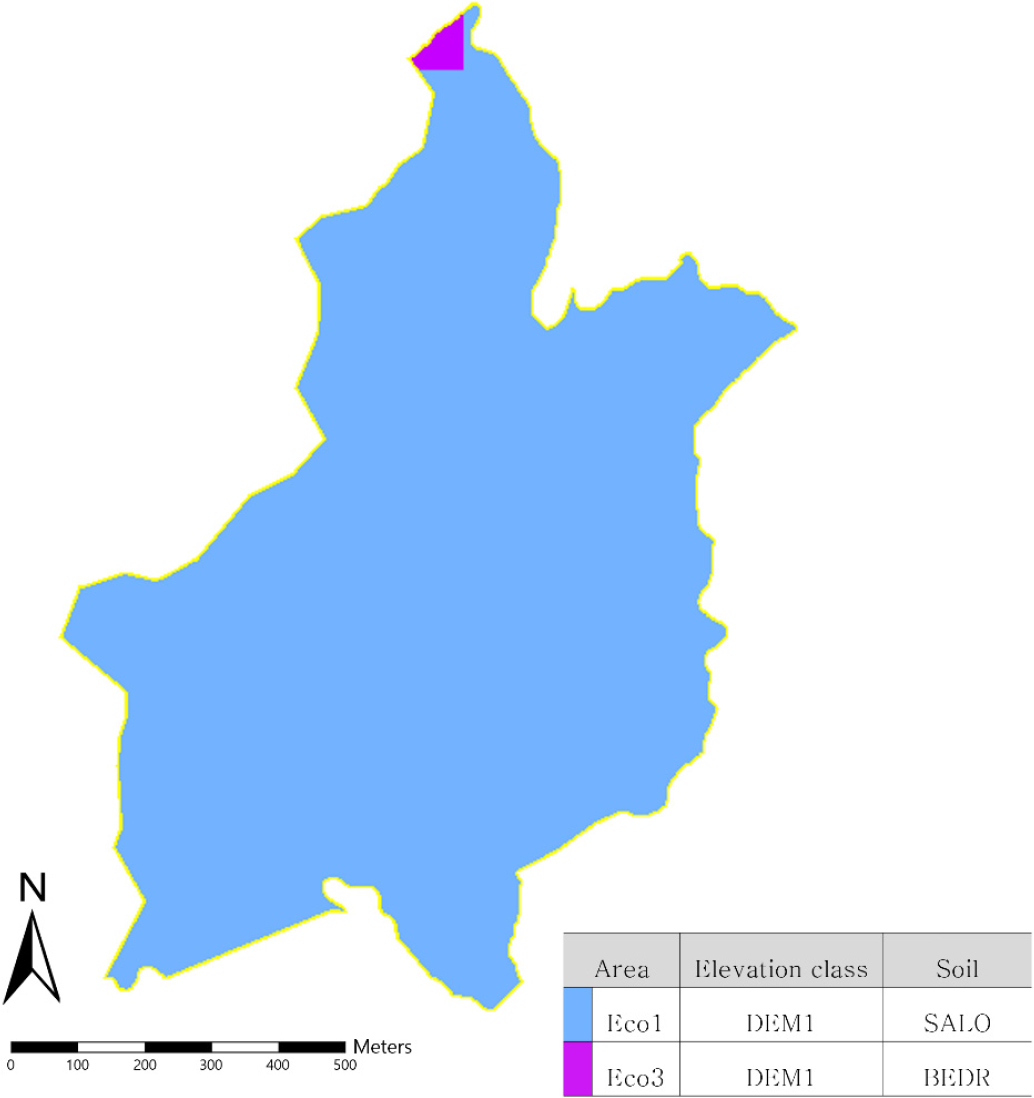
생태역 지도는 지형정보와 그에 따른 환경변수를 적용할 수 있는 핵심 자료이다. 생태역이란, 지역 내 기후, 토양, 지형, 수분 조건 등 생태적 요인이 유사한 공간 단위를 의미하며, 모델 내에서 수종의 생장률, 경쟁력, 물·영양분 순환 등 주요 과정에 영향을 미친다. 이러한 공간적 구분은 경관 내 환경의 이질성을 반영하여, 보다 현실적인 식생천이 과정을 예측할 수 있게 해준다. 생태역 지도 구축을 위해 기온, 지형, 토양 등 주요 환경요인을 활용하였다. 선행 연구에서 연평균 기온으로 생태역을 구분한 것을 반영하여, 기상청의 2000년도 연평균 기온 자료를 활용해 5℃, 9℃, 13℃를 구분 기준으로 설정하였다(Eom, 2019). 지형 정보는 국토지리정보원의 5 m 수치표고모형(digital elevation model, DEM)을 활용하였고 토양은 산림청에서 제작한 산림입지토양도(KFS, 2003)를 참고하였다. 분석 결과, 연구대상지 전체가 연평균 기온 9℃에 해당되었으며, 토양 유형은 사질양토(sandy loam, SALO)와 기타(bedrock, BEDR) 두 가지 유형이 분포하였다. 이러한 환경요인을 통합하여 연구대상지 내 공간을 2개의 유형으로 구분하였다(Fig. 2).
초기 식생지도(initial communities map)
초기 식생지도는 모델 구동의 초기 조건으로서, 각 격자별 수종 구성과 영급 정보를 의미한다(Scheller et al., 2007). 본 연구에서는 1996년 산불 이전의 식생 구성을 기준으로 초기 수종 목록을 설정하여, 산불로 소실된 산림생태계의 자연적 천이 양상을 모의하였다. 산불 이전의 식생 구성을 기준으로 초기 수종 목록을 설정한 이유는, 생태적 기억(ecological memory)이 산불 후 식생천이를 결정한다는 개념에서 기반한 것이다(Johnstone et al., 2016; Spatola et al., 2023). 이는 산불 이전 식생의 종자 생산 능력과 맹아 발달 특성, 그리고 토양 종자은행이 산림의 회복 메커니즘의 핵심 요인으로 작용하기 때문이다(Reed et al., 2025).
초기 식생지도 작성에는 환경처의 제1차 현존식생도(ME, 1990), 산림청의 제3차 수치임상도(KFS, 2003), 산림청의 고성 산불 지역 생태조사 결과보고서(KFS, 1997), 고성군의 고성산불백서(Goseong County, 1997)를 활용하였다. 문헌조사 결과, 산불 이전 우점 수종인 소나무(Pinus densiflora), 신갈나무(Quercus mongolica)였으며 아교목층에는 굴참나무(Quercus variabilis)가 분포하였다. 산불로 인해 모든 식생이 소실되어 천이가 초기 단계에서 시작함을 고려하여, 모든 수종의 영급은 1년으로 설정하였다.
기후자료(climate data)
PnET-Succession 모듈은 수종의 생리·생태 과정을 모사하기 위해 정밀한 월별 기후자료가 요구된다. 본 연구에서는 2000년부터 2024년까지의 기후자료를 구축해 실제 대상지 환경을 반영함으로써 예측의 정확도를 높이고자 하였다.
구축된 기후자료에는 월별 최저/최고 기온, 강수량, 광합성유효복사량(photosynthetically active radiation, PAR), 대기 중 이산화탄소 농도가 포함된다. 월별 최저/최고 기온과 강수량은 기상청의 종관기상관측(automated synoptic observing system, ASOS) 자료 중 연구대상지인 강원도 고성군에 위치한 속초자동기상관측소 관측값을 활용하였다. 식물 생산성의 주요 변수인 광합성유효복사량은 광합성에 이용 가능한 태양 복사 에너지의 범위를 의미하며, 선행 연구에서 제시한 전천일사량 기반 모델 결과로 구축하였다(Choi et al., 2021). 이산화탄소 농도는 연구대상지 장기 관측 자료의 제약으로 인해, 모델의 기본값을 적용하였다.
매개변수(parameters)
매개변수는 수종의 생리·생태적 특성을 정의하는 입력 자료로, 생장 매개변수와 생리 특성(PnET) 매개변수로 구성된다. 수종별 매개변수는 국내 산림생태계 조건을 반영하기 위해 정량적으로 제시된 국내외 선행연구와 LANDIS-II 사용자 가이드의 제시값을 참고하여 설정하였다.
생장 매개변수는 수종의 천이에 대한 반응을 결정하며, 수종의 최대 수명(Longevity), 내음성(Shade Tol.), 내화성(Fire Tol.) 등을 포함한다(Table 2). 생리 특성 매개변수는 잎의 질소 농도(FolN), 광합성이 이루어지는 최저, 최적 온도(PsnT), 광합성 속도를 결정하는 광합성 매개변수(Amax A, B) 등으로 구성된다. 예측의 정확성 향상을 위해 주요 생리 특성 매개변수에 대해 보정 과정을 수행하였다. 각 수종의 임령별 생체량을 산출한 뒤, 예측 결과가 기준값과 일치하도록 생체량 곡선을 단계적으로 조정하였다. 회귀 적합도(R2) 기준을 0.9 이상으로 설정하여 해당 조건을 만족할 때까지 반복 보정을 실시하였다(Table 3). 주요 보정 대상은 잎 포함 질소 함량(FolN), 임관 상층부 특정 잎 무게(SLWmax), 광합성 가능 최소 평균 주간 온도(PsnTMin), 광합성 최적 평균 주간 기온(PsnTOpt), 잎 유지율(FracFol) 등이다(Table 4).
Table 2.
Detailed values for the various parameters by main tree species in this study.
Table 3.
Reference and adjusted aboveground biomass by age classes and main tree species.
Table 4.
Values for PnET Parameters by main tree species.
식생 자료 수집
산불 이후 식생 자료를 수집하기 위해 문헌 조사를 진행하였다. 국립산림과학원에서 발간한 장기 모니터링 보고서(NIFoS, 2007; 2016; 2022)와 기타 선행 연구를 검토해 식생천이 과정과 수종 변화를 분석하였다. 그러나 모니터링이 대상지 내 고정조사구 2곳에 한정되어 연구지역을 대표할 수 없는 한계가 있어 현장 조사를 수행하였다.
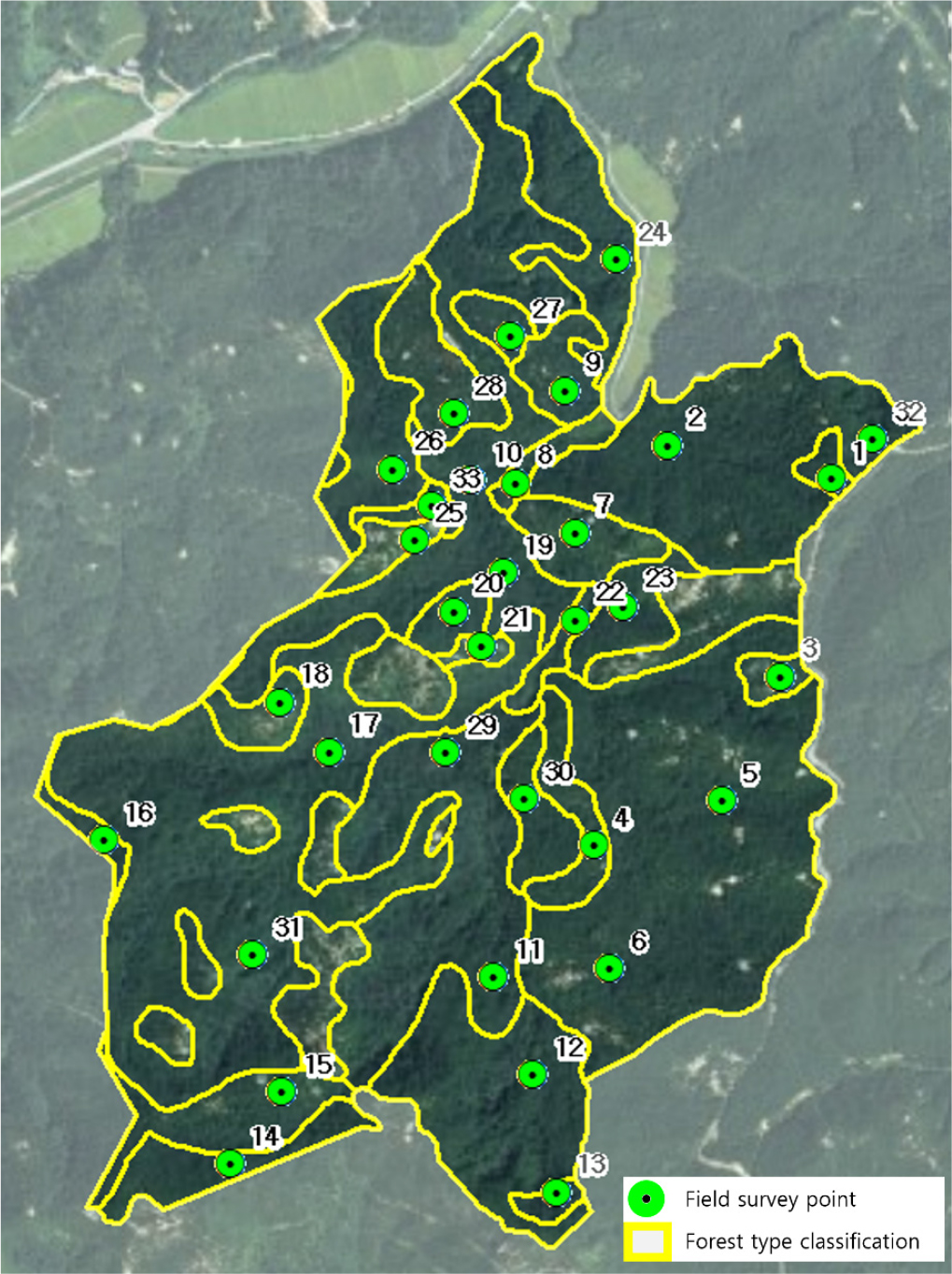
현장 조사 지점은 2023년 산림청에서 배포한 임상도(1 : 5,000)를 기반으로 선정하였다. 동 임상도의 분석 결과, 조사 지역의 식생 분포는 기타 활엽수림 36.1%, 기타 참나무림 33.9%, 소나무림 20.6%, 침활혼효림 4.7%로 나타났으며, 나머지 지역은 암석지 또는 묘지 등으로 구성되어 있었다. 면적이 넓은 지역에서는 두 개 이상의 지점을 설정하여 수종 대표성을 확보하였다. 최종적으로 식생이 존재하지 않는 암석지, 군부대를 제외한 33개 지점을 선정하였으며, 각 지점에서는 교목층과 아교목층을 구분하여 주요 수종 분포를 조사하였다(Fig. 3).
Results and Discussion
LANDIS-II 구동 결과
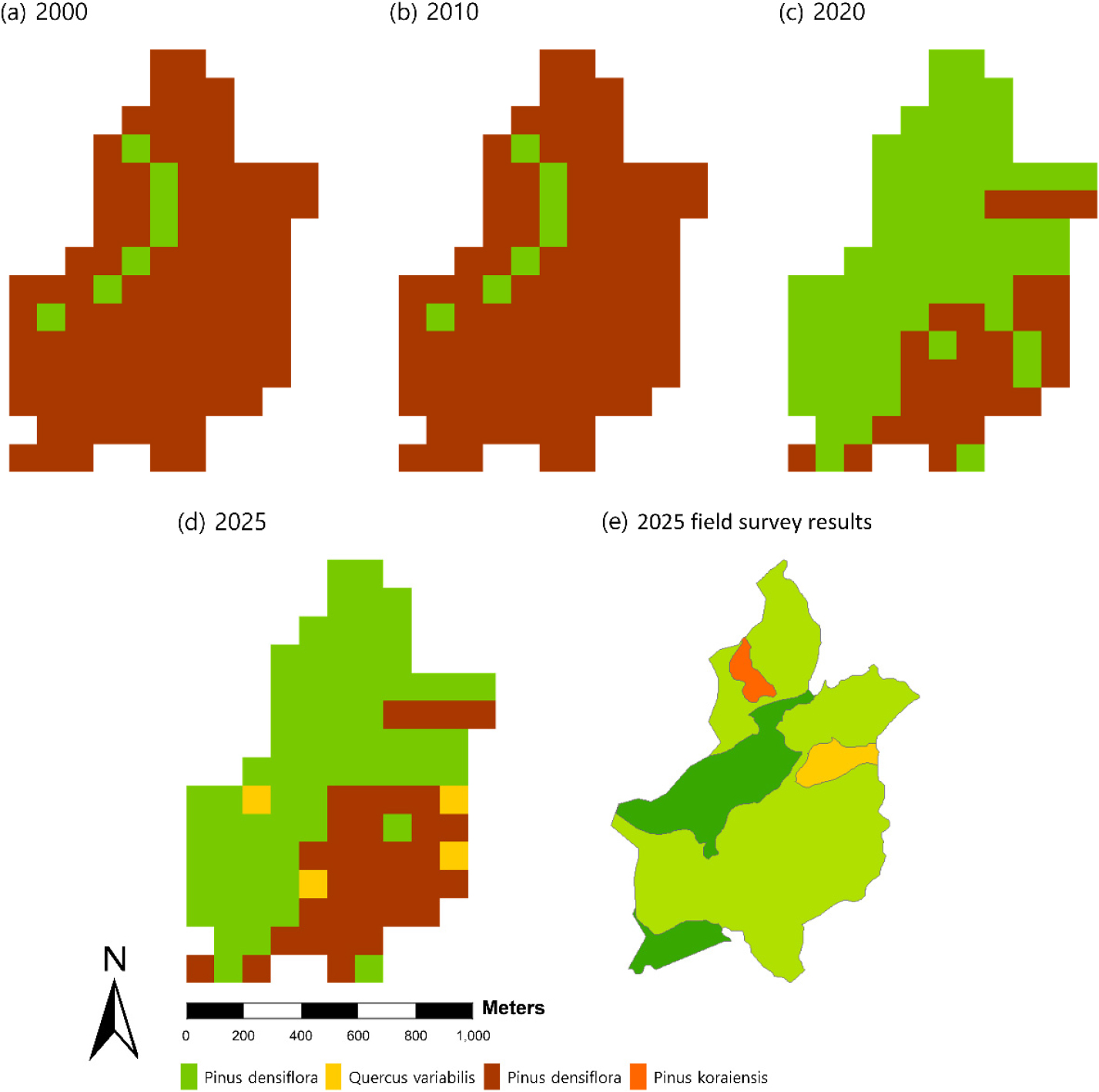
모델 구동 결과, 산불 이후 2000년과 2010년에는 소나무가 전체 면적의 93.3% 분포하며 우세하였고, 신갈나무 분포율은 6.7%에 불과하였다. 그러나 2010년을 기점으로 식생 구조가 전환되어, 2020년에는 신갈나무 70.5%, 소나무 29.5%로 변화하였다. 최종적으로 2025년에는 신갈나무가 전체 면적의 63.8% 분포하였으며, 소나무(32.4%)와 굴참나무(3.8%)가 후속하였다.
모델이 초기 단계인 2000년과 2010년에 소나무 우점을 예측한 것은 소나무의 생태적 특성에서 기인한 것으로 판단된다. 그 근거로 소나무는 높은 광 요구성과 초기 생장률을 바탕으로 개방된 환경과 강한 광 조건에서 경쟁 우위를 확보하며, 건조하고 영양분이 적은 조건에서도 안정적으로 정착할 수 있다(Lee et al., 2004; Choung et al., 2020). 또한, 바람에 의한 원거리 종자 확산 능력이 뛰어나 교란 후 단기간 내에 넓은 면적에 분포할 수 있다는 것이다(Song et al., 2017).
2020년부터 나타난 신갈나무림으로의 급속한 전환은 신갈나무의 장기적 경쟁력에 기인한 것으로 추정된다. 신갈나무는 부분 차광 환경에서도 생장이 가능하며, 긴 수명(약 200 - 400년)과 맹아 재생 능력을 통해 소나무림 하층에서 점진적으로 세력 확장이 가능하다(Song, 2019; Choung et al., 2020). 소나무가 형성한 미세 환경이 개선되면서 신갈나무 치수의 생존률이 증가하였으며, 소나무 수관층 내 개방부가 확대됨에 따라 신갈나무의 상층부 점유가 가속화 된 것으로 판단할 수 있다.
2025년에 나타난 신갈나무의 감소와 소나무의 증가는 모델의 구조적 한계에서 기인한 결과로 해석할 수 있을 것이다. PnET-Succession 모듈은 격자 단위에서 수종과 영급 집단 간의 광·수분 경쟁을 모의하며, 각 집단의 생체량 변화를 독립적으로 계산한다(Gustafson et al., 2023). 모델의 내부 계산 과정에서는 모든 수종의 생체량이 추적되나, 모델 구동 결과물에서는 각 격자 내 여러 수종이 유사한 생체량으로 공존하더라도 지도상에는 가장 높은 단일 수종만이 표현된다(Scheller and Mladenoff, 2007). 따라서 모델 구동 과정에서 미세한 경쟁 관계의 변화로 상대적 생체량 순위 변동이 발생하면, 격자 단위에서는 우점종이 전환된 것처럼 나타난다. 이는 생태계 내에서의 점진적인 경쟁 구도 변화임에도 불구하고, 모델 구동 결과에서는 급격한 식생 교체로 표현되는 현상을 초래한다. 이로 인해 기존 신갈나무로 나타난 격자가 소나무로 변화한 것으로 해석된다.
최종적으로 소나무에서 신갈나무로의 전환은 국내 온대림 산불 피해지에서 일반적으로 보고되는 천이 양상과 일치한다(Lee, 2007; 2022; Choung et al., 2024). 소나무림에서 활엽수 중심의 맹아 재생이 진행되는 자연복원 경향을 모델 구동으로 정량화한 결과, LANDIS-II 모델이 교란 이후 장기적 식생천이 메커니즘을 적절히 반영함을 확인할 수 있었다.
식생 자료 수집 결과
문헌 조사에 따르면 산불 이전 대상지는 소나무림이 우점하였으나(KFS, 1997), 식생 자료 수집 결과 2025년 기준 신갈나무 중심의 활엽수림으로 전환되었다. 조사 지점 33개소의 군락 유형을 분석한 결과, 신갈나무 군락이 전체의 59.7%로 높은 비율을 보였으며, 이어 신갈나무-졸참나무 군락(20.7%), 신갈나무-잣나무 군락(13.5%), 신갈나무-굴참나무 군락(3.5%), 잣나무 군락(2.1%), 신갈나무-산벚나무 군락(0.5%) 순으로 나타났다. 현장 내 지점별 분포 양상은 인공조림지인 잣나무 군락으로 제외하고 신갈나무 중심으로 유사하게 나타났다. 소나무는 인근 인공복원지 인접부 혹은 계곡부에서 일부 확인되었으나, 전체 지역에서는 우점종으로서의 지위를 상실한 것으로 해석된다.
산불 이전 우점 수종인 소나무의 쇠퇴는 화재 취약성에서 기인하는 것으로 판단된다. 소나무는 산림 하층에서 발생하는 지표화에 취약한 수종으로, 수피가 얇아 열손상을 받기 쉽고 산불 발생 시 고사하는 경우가 대다수이다(Kwon et al., 2021). 1996년 대상지에 피해를 입힌 고성 산불은 지표화였으며(Goseong County, 1997; KFS, 1997), 이로 인해 성목 개체군이 파괴되었다. 이후 산불로부터 회복되는 과정에서 소나무는 활엽수와의 경쟁에서 밀려나며 우점종으로서의 지위를 상실한 것으로 해석할 수 있을 것이다.
소나무의 쇠퇴와 대조적으로 신갈나무를 포함한 참나무류가 우위를 차지한 것은 산불 이후 활엽수종의 맹아 재생 능력에서 비롯한다. 참나무류는 산불로 인해 지상부가 소실되더라도, 잔존하는 지하부 뿌리에 저장된 에너지원을 활용하여 맹아 발생이 가능하다(Choung et al., 2020). 이러한 재생 능력을 통해 참나무류는 산불 직후 빠르게 회복하며, 광 환경이 개선된 조건에서 급속한 생장을 보인다. 결과적으로 소나무 우점림이었던 대상지에서 활엽수종이 우위를 점하는 새로운 천이 궤도가 형성된 것으로 예상된다.
연구대상지 식생 변화 비교 분석
연구대상지의 2025년 식생천이 과정을 LANDIS-II 구동 결과와 현장 조사 자료를 우점 수종 기준으로 비교하였다(Fig. 4).
모델 구동 결과, 신갈나무림은 전체 면적의 63.8%, 굴참나무림은 3.8% 나타나, 신갈나무 중심의 활엽수림 우점 경향이 예측되었다. 현장 조사에서는 신갈나무림 73.7%, 졸참나무림 20.7%, 굴참나무림 3.5%로 나타나 모델과 유사한 천이 양상을 보였다. 이러한 결과는 LANDIS-II 모델이 교란 이후 산림생태계 구조 변화를 안정적으로 모사할 수 있음을 시사한다(Stevens-Rumann and Morgan, 2016).
모델과 현장 간 차이는 소나무 분포율에서 나타났다. 모델 구동 결과에서는 32.4%로 예측되었으나, 현장에서는 일부 지역에 제한적으로 분포하며 우점종으로의 지위를 상실하였다. 이러한 차이는 두 가지 요인으로 설명할 수 있다.
첫째, 매개변수의 불확실성이다. PnET-Succession 모듈은 수종별 생장과 생존을 모의하기 위해 총 28개의 생리적 매개변수를 필요로 하며, 입력값의 정확성에 따라 결과에 영향을 미친다(McKenzie et al., 2019; Gustafson et al., 2023). 본 연구에서는 국내외 선행 연구 자료와 모델 기본값을 바탕으로 수종별 매개변수 보정을 수행하였으나, 주요 생리 매개변수 6개에 제한하여 보정을 수행했으며, 국내 자료가 부재한 매개변수는 모델 기본값을 사용하였다. 이로 인해 소나무를 비롯한 주요 수종의 실제 생리·생태적 특성을 충분히 반영하지 못한 것으로 사료된다(Remy et al., 2019).
둘째, 연구대상지 규모의 한계이다. 연구대상지 면적은 82 ha로, 모델 구동 면적은 인근 지역을 포함하여 105 ha이다. 그러나 LANDIS-II는 10,000 ha 이상의 경관 단위를 대상으로 개발된 모델로, 상대적으로 작은 규모는 경계 효과로 인한 외부 교란 유입을 놓치게 되어 예측 정확도를 저하시킨다(Keane et al., 2002). 소규모 경관에서는 모델 경계 외부에서 유입되는 종자 산포 등의 외부 교란이 충분히 반영되기 어렵다(San-José et al., 2022; Arroyo-Rodríguez et al., 2023). 이러한 한계로 인해 식생천이의 양상은 재현 가능하였으나, 구체적인 수종 분포 비율에는 차이가 생긴 것으로 해석된다.
Conclusion
본 연구는 1996년 고성 산불과 2000년 동해안 산불 피해지를 대상으로 LANDIS-II 모델과 PnET-Succession 모듈을 활용하여 25년간의 식생천이 과정을 정량적으로 예측하고 현장 조사 자료와의 비교를 통해 모델의 국내 적용 가능성과 한계를 분석하여 생태적 산림 복원 계획 수립을 위한 기초자료를 제공하고자 하였다.
연구 결과, 모델과 현장 조사 모두 신갈나무 중심 활엽수림 우점을 확인하여(모델 구동 63.8%, 현장 73.7%), 모델이 산불 이후 침엽수에서 활엽수로의 천이 방향성을 재현할 수 있음을 증명하였다. 그러나 소나무 분포율은 32.4%인 모델과 우점종 지위를 상실한 현장 간 차이를 보였다.
본 연구의 결과는 다음의 한계를 가진다. 첫째, 생리 특성 매개변수 28개 중 6개만을 보정하여 적용하였다. 둘째, 대상지 규모(82 ha)가 LANDIS-II 권장 규모(10,000 ha 이상)보다 작아 경계 효과, 종자 유입 등의 요인 반영에 제약이 있었다. 이러한 한계점은 향후 개선이 필요하나, 산불 피해지의 자연복원 과정을 정량적으로 예측할 수 있는 방법론적 가능성을 보였으며, 국내 수종 매개변수 보정의 정교화 필요성을 제시하였다. 이는 산림 복원 계획 수립 시 장기 천이 과정 예측을 위한 기초자료로 활용 가능할 것으로 기대된다. 향후 연구에서는 국내 주요 수종의 생리 매개변수를 체계적으로 구축하여 모델 정확도를 향상시켜야 하며, 넓은 면적에서 모델을 적용하여 경관 수준의 천이 과정을 분석하여야 한다. 또한, 대상지가 아닌 산불 피해지에 적용하여 지역별 천이 특성을 비교하고 모델의 일반화 가능성을 검증할 필요가 있다.
본 연구는 LANDIS-II 모델이 국내 산불 피해지의 장기 식생천이를 재현 가능하나, 매개변수 정확성, 공간 규모 등 일부 측면에서 지속적인 개선의 여지가 있음을 제시하였다. 이를 바탕으로 모델을 활용한 산불 피해지 식생천이 분석 방법론을 제안하고, 자연복원지의 장기 모니터링 자료와 예측 결과를 통합적으로 해석하는 접근을 시도하였다. 또한 국내 주요 수종의 생리·생태적 특성에 대한 매개변수 구축과 보정 절차는 향후 유사 연구의 기초 자료로 활용될 수 있을 것이다. 나아가 기후변화 및 교란 시나리오를 적용한다면, 변화하는 환경 조건에서 식생천이의 과정을 정량적으로 비교·평가할 수 있을 것으로 기대된다.
